ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 2 - Г.Й. Щербак - 1996
ТИП ЧЛЕНИСТОНОГІ (ARTHROFODA)
ПІДТИП ЗЯБРОДИШНІ, АБО РАКОПОДІБНІ (BRANCHIATA, АБО CRUSTACEA)
До цього підтипу належать організми, що найбільш повно освоїли водне середовище: вони зустрічаються в пересихаючих калюжах, прісних і морських водоймах, заселюючи всю товщу води, від глибоководних морських западин до гіпонейстона (поверхневої плівки води). Більшість із них — вільноживучі, рухливі тварини, серед них є й сидячі, прикріплені до субстрату види, а також паразити інших водяних тварин. Невелика група ракоподібних перейшла до наземного існування. Описано понад 40 тис. видів.

Розміри ракоподібних коливаються від часток міліметра до 80 см. Для ракоподібних характерна велика різноманітність у зовнішній будові тіла й кінцівок, тоді як внутрішня організація досить однотипна, хоч у представників різних класів є певні відмінності. Загальну схему будови ракоподібних показано на рис. 50.
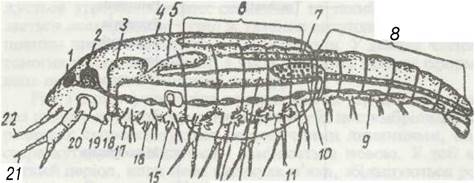
Рис. 50. Схема організації ракоподібних
Тіло ракоподібних сегментоване. Кількість сегментів коливається в широких межах. Тіло поділене на три тагми — головну, грудну та черевну. Сегменти кожної тагми можуть бути чітко відмежовані один від одного або так чи інакше злиті. Іноді сегменти грудного відділу на черевній стороні чітко відмежовані, а на спинній утворюється загальний спинний щит (карапакс), що прикриває частину тіла у вигляді пласкої або двосхилої покрівлі. Між окремими тагмами сегменти розподіляються так.
До складу голови входять акрон і кілька сегментів. Сегментарний склад голови ракоподібних, як і інших членистоногих, різними дослідниками визначається в чотири—вісім сегментів, тому й належність її придатків до того чи іншого сегмента дискусійна. Наприклад, перші антени одні вважають придатками акрону, інші — I, II або навіть III сегмента.
Груди і черевце представників більшості класів ракоподібних мають різну кількість сегментів, крім видів класу Маlacostraca, в яких до складу грудної тагми постійно входять вісім, а черевної — шість-сім сегментів. Черевце, як правило, закінчується анальною лопаттю (тельсоном), на якій часто є пара придатків (фурка, або вилка).
Часто поділ на три тагми порушується. Наприклад, іноді до складу голови входить лише її частина, що несе антепули та антени, тоді ми говоримо про протоцефалон. При цьому щелепні сегменти можуть залишатися вільними або об'єднуватися з грудними, утворюючи нову тагму — щелепогруди (гнатоторакс). Навпаки, у багатьох форм процес цефалізації
охоплює не тільки сегменти голови, а й один-два передні грудні сегменти, утворюється тагма, що зветься сиицефалоном. Інколи голова й усі грудні сегменти зливаються в головогруди, або цефалоторакс.
У деяких ракоподібних кілька задніх сегментів черевця зливаються з тельсоном, утворюючи так званий плеотельсоп,
Кінцівки різних тагм і сегментів тіла мають різну будову та функції, та все ж можна охарактеризувати загальний план їх будови (рис. 51). Кожна кінцівка складається з основної частини — протоподита, що має один—три членики, від неї відходить двогілляста частина — телоподит. Зовнішня гілка — екзоподит та внутрішня — ендоподит мають різну, часто велику, кількість члеників. У багатьох ракоподібних на певних сегментах тіла від протоподита відходять також один-два дихальні відростки — епіподити, що виконують функцію шкірних зябер.
Кінцівки голови представлені п'ятьма парами. Перші з них — антенули (антени Т), як правило, одногіллясті, йлише у представників класу Malacostraca розщеплюються на дві й навіть три гілки. Найчастіше антенули функціонують як органи чуття (дотику, нюху), хоч іноді можуть брати на себе й плавальну функцію або ставати органами прикріплення. Друга пара кінцівок — антени (антени II) — типово двогіллясті, але нерідко одна з гілок редукується. Так, у всім відомого річкового рака ендоподит антен утворює довгий членистий «бич», тоді як екзоподит має вигляд короткої пластинки. Третя пара кінцівок — жувальця, або мандибули, відіграє головну роль при подрібненні їжі. Мандибули тільки у личинок (наупліус) двогіллясті; у більшості дорослих форм телоподит майже повністю зникає, протоподит утворює жуйну лопать і перетворюється на масивну зазубрену верхню щелепу. Наступні дві пари кінцівок — нижні щелепи, або макети, — багато членисті, з ніжним телоподитом; членики протоподиту мають спеціальні жуйні відростки.
Грудні кінцівки або всі однакові, або передні (1—3 пари) видозмінюються в ногощелепи. Вони втрачають рухову функцію й беруть участь у захопленні та подрібненні їжі. Грудні кінцівки часто бувають двогіллястими, але нерідко одна з гілок (як звичайно, екзоподит) цілком або частково редукується. Функція грудних кінцівок здебільшого локомоторна (плавальна, ходильна ) або хапальна, дихальна тощо.
Черевні кінцівки є тільки у Malacostraca. Вони, як правило, двогіллясті; виконують найчастіше не рухову, а інші функції: дихальну, органів копуляції тощо, але остання пара черевних кінцівок багатьох десятиногих раків перетворюється на потужні плавальні пластинки.
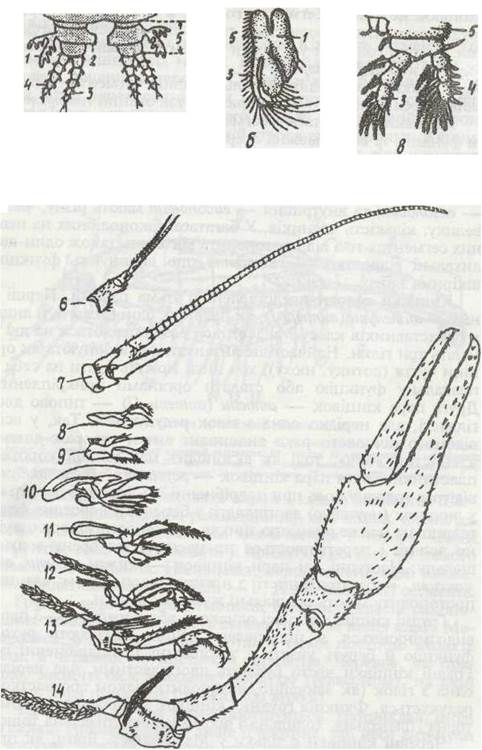
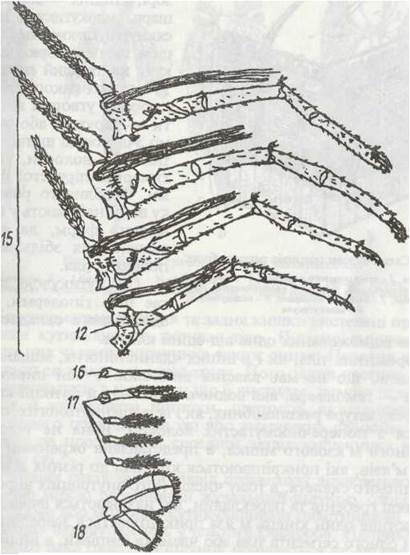
Рис. 51. Кінцівки ракоподібних
У деяких паразитичних раків кінцівки втрачають членистість або, в дорослому стані, зовсім зникають (Sacculina).
Зовні тіло ракоподібних укрите кутикулою, в якій, на відміну від інших членистоногих, відсутній водонепроникний шар епікутикули, й тому на суші через їхні покриви вільно випаровується вода. Пігменти, що обумовлюють забарвлення, розташовані у відносно тоненькій екзокутикулі. Ендокутикула порівняно товста (рис. 52). У багатьох дрібних ракоподібних покриви порівняно м'які й прозорі; в інших —зовнішні шари ендокутикули просякнуті вуглекислим кальцієм, що перетворює кутикулу на твердий панцир. У багатьох ракоподібних кутикула утворює вирости — нерухомі або рухомо зчленовані шипи, щетинки та волоски, часто химерної пірчастої форми. Особливого розвитку вони набувають у плаваючих форм, де призначені для збільшення поверхні тіла.
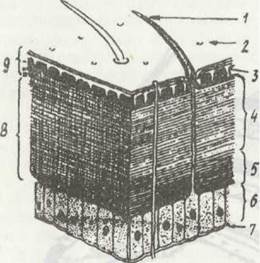
Рис. 52. Схема будови покривів ракоподібних:
1 - щетинка; 2 - отвір перового канальця; 3 - пігментний шар; 4 - вапняковий шар; 5 — невапняковий шар; 6 - гіподерма; 7 - базальна мембрана; 8 - ендокутикула; 9 - екзокутикула
Під кутикулою залягає шар гіподерми, яка здебільшого складається з добре відмежованих одна від одної клітин.
Порожнина тіла, як і в інших членистоногих, мішана — міксоцель, що не має власної вистилки. У ній циркулює рідина — гемолімфа, яка водночас виконує й функції крові.
Мускулатура ракоподібних, як і всіх членистоногих, складається з поперечносмугастих волокон. Вона не утворює суцільного м'язового мішка, а представлена окремими групами м'язів, які прикріплюються кінцями до різних ділянок зовнішнього скелета, в тому числі і його внутрішніх виростів у вигляді гребенів та перекладин, які називаються апофізами. Найчастіше один кінець м'яза прикріплюється зсередини до стінки одного сегмента тіла або членика кінцівки, а інший — до стінки другого. Особливо потужні м'язи підходять до мандибул. У форм із двостулковим карапаксом є особливий м'яз-замикач, що тягнеться від однієї стулки до іншої впоперек тіла.
Травна система (рис. 53) складається з передньої, середньої та задньої кишок. Рот міститься на черевній стороні голови; спереду і ззаду він обмежений непарними кутикулярними складками — верхньою та нижньою губами, а по обидва боки від нього розташовані мандибули і махсили, призначені для подрібнення їжі.
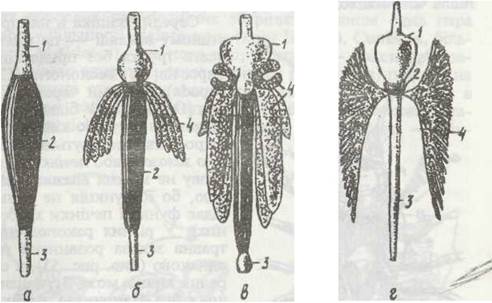
Рис. 53. Будова травної системи ракоподібних:
а - Copepoda; б - Leptostraca; в - Euphausiacoa; г - Decapods; 1 - передня кишка, що іноді розширюється в жувальний шлунок; 2 - середня кишка; 3 - задня кишка; 4 -травна залоза (печінка)
Ектодермальні передня та задня кишки вистелені продовженням кутикули, що вкриває тіло. Під час линяння кутикула цих частин кишечника також линяє. Передня кишка складається з довгого стравоходу. Його задня частина може розширюватись, а кутикула, що її вистилає, значно потовщуватися й утворювати шипики, щетинки, які служать для додаткового подрібнення їжі або виконують функцію фільтра, що не пропускає в середній відділ кишечника великі шматки їжі.
У вищих раків задній відділ стравоходу відокремлюється в різною мірою розвинений жувальний шлунок, озброєний щетинками або міцними кутикулярними пластинками. Найскладнішу будову він має у донних Decapoda. Наприклад, у річкового рака (рис. 54) шлунок поділяється на дві частини: кардіальну (вхідну) та пілоричну. Кардіальна частина служить для додаткового подрібнення їжі. Вона вистелена товстою кутикулою, яка особливо розвинена в трьох місцях, утворюючи жувальні пластинки з гострими зубцями («шлунковий млин»), Пілоричний відділ має складнішу будову: тут є так званий прес, стінки якого мають добре розвинеш м'язи; в ньому подрібнена їжа спресовується, і з неї віджимаються рідкі фракції. Великі частинки їжі, що не подрібнилися щелепами і «шлунковим млином», потрапляють безпосередню в задню кишку й виводяться назовні. Рідкі фракції їжі ще раз проціджуються за допомогою довгих кутикулярних волосків, після чого надходять до коротенької середньої кишки.
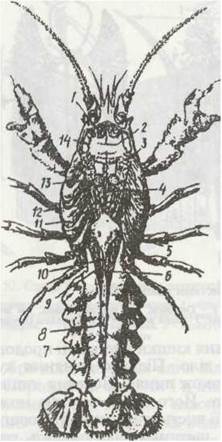
Рис. 54. Внутрішня будова самця річкового рака:
1 - передні м'язи шлунка; 2 ~ шлунок; 3 - м'язи мандибул; 4 - аорта; 5 - серце; 6 - задня дорзальна артерія; 7 - м'язи черевця; 8 - задня кишка; 9 - сім'япровід; 10 - сім'яник; 11- зябра; 12 - травна залоза (печінка); 13 -задні м'язи шлунка; 14 -антенальна залоза
Середня кишка в найпростішому вигляді — це пряма довга трубка без придатків і виростів, як у веслоногих (Соpepoda), частини черепашкових (Ostracoda). У більшості ж вона має одну або кілька пар відростків, що звуться травною залозою, або печінкою (цю назву не можна вважати вдалою, бо її функція не відповідає функції печінки хребетних). У різних ракоподібних травна залоза розвинена неоднаково (див. рис. 53), а середня кишка може бути різної довжини. Наприклад, у річкового рака її довжина становить приблизно 1/20 довжини всього кишечника. Залозисті клітини печінки виділяють ферменти, які розщеплюють жири, білки та вуглеводи. Тут же відбувається і всмоктування. Внутрішньоклітинне травлення в печінці не відбувається.
Задня кишка, як правило, коротка. Лише в деяких видів вона досягає значних розмірів.
У деяких паразитичних раків (Sacculina) травна система редукована. Вони всмоктують соки хазяїна всією поверхнею тіла.
Живляться ракоподібні різною їжею. Одні фільтрують з води органічні рештки та дрібні організми — бактерії, одноклітинні водорості, різні безхребетні; інші активно відривають жувальцями шматки живих або мертвих тварин і рослин.
Видільна система раків майже втратила метамерний характер. Вони мають дві пари видільних органів, які розташовані біля основ антен (антенальні залози) та другої пари нижніх щелеп (максилярні залози). Протягом життя у ракоподібних звичайно розвішаються обидві пари видільних органів, однак дуже рідко (дорослі морські Ostmcoda, Leptostгаса, деякі Mysidacea) вони функціонують одночасно. В інших дорослих ракоподібних зберігається лише одна пара залоз: у більшості Malacostraca (крім Isopoda, Cumacea, більшості Mysidacea) — антенальні, у решти — максилярні залози. Часто одна пара залоз (у різних раків різна) працює на личинкових стадіях, зношується і замінюється на іншу в дорослому віці. Наприклад, у Copepoda на личинкових стадіях функціонують антенальні залози, а в дорослих — максилярні.
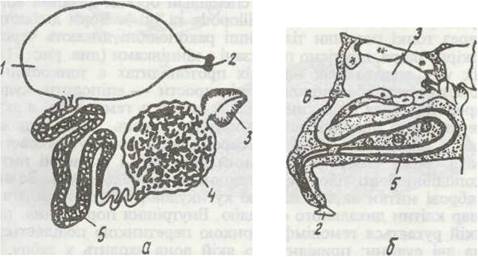
Рис. 55. Видільна система ракоподібних:
а-схема будови антенальиої залози Astacus; б - поперечний зріз максилярної залози Cypris; 1 - сечовий міхур; 2 - видільний отвір;
3 - целомічний мішечок; 4 -лабіринт; 5 - нефридіальний канал; 6 — клапан
У типовому випадку видільна залоза має кінцевий мішечок (залишок целома) та вивідний канал (видозмінений целомодукт), який може ускладнюватися (рис. 55). Так, у більшості Decapoda він складається з лабіринта і сечового міхура, а в річкового рака лабіринт має дві частини: власне лабіринт і нефридіальний канал. Внутрішні стінки лабіринта утворюють численні складки й вирости в просвіт каналу, які значно збільшують його поверхню. У різних ділянках вивідного каналу відбуваються процеси зворотного всисання (реабсорбція) води, мінеральних солей та деяких органічних речовин.
Основним продуктом розкладу азотовмісних речовин у ракоподібних, як водяних тварин, є аміак.
Видільні залози ракоподібних одночасно є й органами осморегуляції. Деякі ракоподібні здатні переносити різкі коливання солоності (евригалинні організми). Так, прибережні краби можуть зносити опріснення води під час злив; деякі бокоплави заходять у гирла річок, сакуліна живе у воді різної солоності тощо. Одним з механізмів, що забезпечує цю здатність, є робота їхніх видільних органів. В опріснених водах об'єм сечі в них збільшується, а концентрація солей у ній зменшується, чим забезпечується стабільна концентрація іонів у гемолімфі.
У виділенні бере участь також жирове тіло, побудоване приблизно так само, як і в інших членистоногих (детально розглядається на прикладі комах у відповідному розділі підручника).
У багатьох ракоподібних спеціальні органи дихання відсутні (класи Ostmcoda, Maxillopoda та ін. ). Вони дихають через тонкі покриви тіла. Інші ракоподібні дихають через шкірні зябра, що тісно пов'язані з кінцівками (див. рис. 51). Як уже згадувалося, на їхніх протоподитах є тонкостінні пластинчасті або мішкоподібні вирости — егаподити, куди продовжується порожнина тіла й заходить гемолімфа, в яку через ніжні покриви зябер дифундує кисень. Часто будова зябер ускладнена. Так, у Decapoda, наприклад у річкового рака, зябра мають центральний стрижень і численні ниткоподібні бічні гілочки, розташовані навколо нього. Зовні зяброві нитки вкриті тонкою кутикулою, під якою залягає шар клітин дихального епітелію. Внутрішня порожнина, по якій рухається гемолімфа, тонкою перетинкою поділяється на дві судини: привідну, по якій вона входить у зябру, і вивідну, по якій виходить. Зябра у річкового рака містяться під бічними складками головогрудного щита, у двох зябрових порожнинах. У кожній із них зябра розташовані трьома поздовжніми рядами. Зябра одного ряду сидять на протоподитах ніг, другого — там, де з'єднуються протоподити з тілом, третього — на бічній стінці тіла. Вода через зяброву порожнину (спереду назад або навпаки) рухається спеціальними відростками другої пари нижніх щелеп, які коливаються до 200 разів за хвилину.
Цікаві зміни дихального апарата у деяких раків, які перейшли до наземного життя. Так, у краба-розбійника (Birgus latro), що живе на суші, на островах Тихого океану, на внутрішніх стінках зябрових порожнин з'являються численні гроноподібні складки шкіри, які мають щільну сітку кровоносних судин, тоді як зябра редукуються. Таким чином зяброві порожнини перетворюються на своєрідні легеневі порожнини, і дихання стає повітряним.
У Isopoda дихання пов'язане не з груд ними, як у більшості ракоподібних, а з черевними кінцівками. У них на протоподитах передніх п'яти сегментів черевця Є широкі листоподібні тонкостінні вирости, що накладаються один на одного, як сторінки книги. Одна пара черевних ніжок звичайно сильно склеротизована, вона покриває всі інші як кришечка. Гака будова дихального апарата, мабуть, їдала змогу частині рівноногих перейти до сухопутного існування, оскільки зябра в них виявилися добре захищеними від висихання. У частини наземних Isopoda, наприклад у мокриць, на пластинчастих черевних ногах є глибокі розгалужені вп'ячування покривів. Порожнина кінцівки заповнена гемолімфою, яка омиває ці вп'ячування. Всередину порожнини через спеціальний отвір (дихальце) заходить повітря, звідки кисень Дифундує в гемолімфу. Це трахейні легені (рис. 56). Вони дуже нагадують трахеї комах.
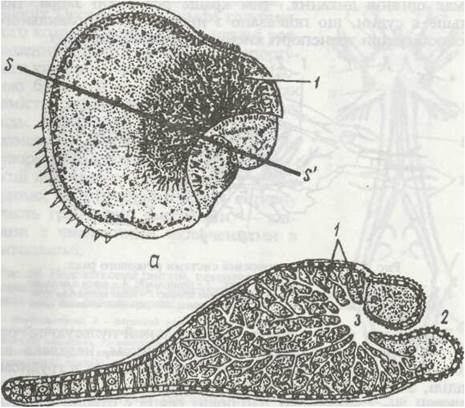
Рис. 56. Дихальні черевні ніжки мокриці Porcellio scaber:
а - зовнішній вигляд; б - зріз по лінії s-f. 1 - псевдотрахеї; 2 - дихальце; 3 - повітряна порожнина
Кровоносна система ракоподібних незамкнена, тобто гемолімфа тече не тільки по судинах, а й виливається в міксоцель (рис. 57). Будова кровоносної системи певною мірою залежить від ступеня розвитку органів дихання. Вона найбільш редукована або зовсім відсутня у ракоподібних, у яких немає органів дихання. Чим краще розвинеш зябра, тим більше є судин, що пов’язано з необхідністю правильного кровообігу при транспорті кисню від органів дихання.
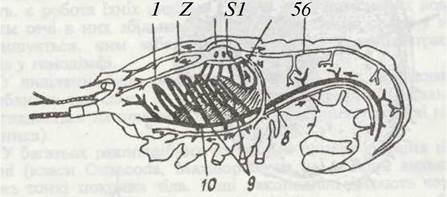
Рис. 57. Схема кровоносної системи річкового рака:
1 - антенальна артерія; 2 - передня аорта; 3 - перикардій; 4 - серце з остіями;
5 - зяброво-серцеві канали; 6 —задня дорзальна артерія; 7 -задня вентральна артерія; 8 - низхідна артерія; 9 - зяброві судини; 10 - черевний венозний синус
Звичайно у ракоподібних є центральний пульсуючий орган — серце, що лежить над кишечником, недалеко від зябер. У більшості ракоподібних воно міститься в грудному відділі, а в Isopoda, в яких зябра зв'язані з черевними кінцівками, — у черевному. По боках серця є отвори з клапанами (ости), через які кров надходить до серця. У зяброногих, ротоногих раків та бокоплавів серце має вигляд багатокамерної трубки з багатьма парами остій, тоді як у десятиногих раків — це короткий мішечок із трьома парами остій, а в гіллястовусих — з однією. Від серця майже завжди відходить передам і рідше задня аорта; від аорт можуть відходити додаткові артерії. Врешті-решт гемолімфа з артерій потрапляє в порожнину тіла, звідки по її відокремлених ділянках (синусах) іде до зябер, де збагачується киснем і по спеціальних каналах, що є відділами міксоцеля («зяброво- ерцеві канали»), протікає в ділянку міксоцеля, що оточує серце — перикардій, а потім, через остії, в саме серце. Усі згадані ділянки міксоцеля (синуси, перикардій) відокремлені тоненькими плівками, які утворюються з проміжної речовини і не мають клітинної структури.
Отже, у ракоподібних є лише артерії, що несуть кров від серця, але відсутні вени; до серця кров рухається по ділянках міксоцеля. Крім того, в них нема капілярів.
До складу гемолімфи входять досить різноманітні клітини, переважно амебоїдної форми, здатні до фагоцитозу. У гемолімфі присутні речовини, що вбивають патогенні мікроорганізми, однак білки типу імуноглобулінів хребетних, з яких утворюються антитіла, відсутні.
Кисень розчиняється в плазмі; у деяких Decapoda гемолімфа голубуватого кольору, оскільки в її плазмі розчинений пігмент, здатний транспортувати кисень — гемоціанін. Він за будовою близький до гемоглобіну, однак замість заліза містить двовалентну мідь. Гемолімфа має здатність згортатися при пошкодженні покривів та утворювати згусток; механізм цієї реакції досі вивчено недостатньо, однак відомо, що в згортанні беруть активну участь гранулярні амебоцити — клітини з численними включеннями в цитоплазмі.
Нервова система ракоподібних (рис. 58) — це парний надглотковий ганглій, або головний мозок, навкологлоткові конективи та пара черевних нервових стовбурів із парними гангліями в кожному сегменті. Надглотковий ганглій складається з переднього мозку, або протоцеребрума, який іннервує очі, середнього — дейтоцеребрума, від якого відходять нерви до антенул, та заднього — тритоцеребрума. Нерви до антен II відходять від тритоцеребрума або навкологложових конектив.
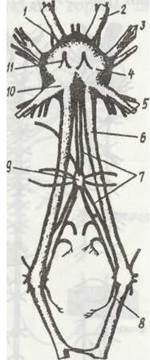
Рис. 58. Надглотковий ганглій, навкологлоткові конективи та вегетативна (стоматогастрична) нервова система річкового рака з черевної сторони: 1 - протоцеребрум; 2 - оптичний нерв; 3 - нерви рецепторів головогрудей; 4 - дейтоцеребрум; 5 - нерви антен; 6 - навкологлоткова конектива; 7 - вегетативна (стоматогастрична )нервова система; 8 - конективний ганглій; 9 - глотковий
(стравохідний) ганглій; 10 - тритоцеребрум; 11 - нерв антен
Найпримітивнішу нервову систему мають зяброногі та цефалокариди. У них черевні стовбури широко розставлені, а парні ганглії сполучаються довгими подвійними поперечними комісарами (рис. 59, а). У більшості ракоподібних обидва ганглії одного сегмента зближені, поперечна комісура між ними не помітна, хоча конективи часто залишаються розділеними. У багатьох ракоподібних відбувається повне злиття не тільки парних гангліїв, а й конектив у один непарний нервовий ланцюжок (рис. 59, б—г). Крім того, відповідаю до злиття окремих сегментів тіла спостерігається вкорочення черевного нервового ланцюжка і злиття гангліїв сусідніх сегментів. Так, у річкового рака ганглії, що іннервують щелепи та ногощелепи, утворюють великий підглотковий вузол, а у крабів взагалі всі черевні ганглії зливаються в один (рис. 59, д, є).

Рис. 59. Будова нервової системи ракоподібних:
а - Anostraca; б - Euphausiacea; в - Stomatopoda; г, д - Decapoda; e - Copepoda; є — Ostracoda
Ганглії черевного нервового ланцюжка або драбини іннервують органи відповідного сегмента (рецептори, м'язи кінцівок), при їх злитті залишаються нерви, які тягнуться від складного ганглія до відповідного сегмента.
До гангліїв нервової системи, поряд із нервовими, входять групи нейросекреторних клітин, розташованих у протоцеребрумі, тритоцеребрумі та в гангліях черевного нервового линяння — екдистерон. Крім линяння, екдистерон стимулює також інтенсивність обміну речовин і ріст. Крім того, Y-залоза продукує гормон, який стимулює розвиток статевих залоз.
У зоні очного стебельця або поблизу основи очей є група нейросекрегорних клітин (Х-орган), стероїдні гормони яких потрапляють у спеціальний резервуар — синусну залозу, а звідти — в гемолімфу. Ці гормони є антагоністами екдистерону й запобігають линянню.
Y-залоза функціонує лише у статевонезрілих особин; у дорослих її діяльність припиняється, інколи вона зовсім атрофується. Навпаки, Х-залоза починає функціонувати на заключних етапах онтогенезу і не припиняє секреції аж до смерті тварини.
У стінках проток сім'яників є андрогенна залоза, гормони якої стимулюють розвиток сім'яників і вторинних чоловічих ознак
ланцюжка. Роль нейросекретів вивчено недостатньо. У ракоподібних, крім цього, є й спеціальні залози внутрішньої секреції. У голові є так звані Y-органи. У більшості ракоподібних вони знаходяться в антенальному, а у Malacostraca — у другому максилярному сегментах (рис. 60). Ці органи мають різну форму (лисгкоподібну, конічну тощо).
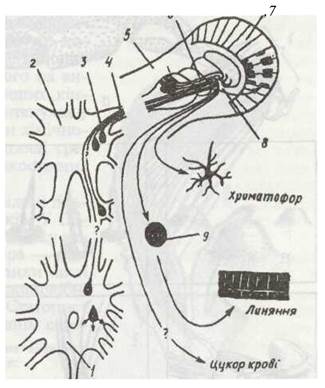
Рис. 60. Схема нейросекреторної системи синусової залози краба:
1 - черевна гангліозна маса; 2 - головний мозок; 3 - нейросекреторні клітини; 4 - зоровий нерв; 5 - зорове стебельце; б — відростки не йро секреторної клітини; 7 - складне око; 8 - синусова залоза; 9 - ендокринна залоза
Гормон Х-залози впливає також на забарвлення тіла. У гіподермі багатьох крабів, креветок тощо є спеціальні зірчасті клітини-хроматофори. У них містяться гранули пігментів різного кольору. Коли, наприклад, гранули червоного кольору рівномірно розсіяні в клітині, він помітний, коли ж вони концентруються в центрі, колір зникає. Під впливом нервових імпульсів від очей X-залоза виділяє гормон, що впливає на розподіл певного пігменту (всього відомо чотири таких гормони), і рак змінює колір (захисне, загрозливе чи інше забарвлення).
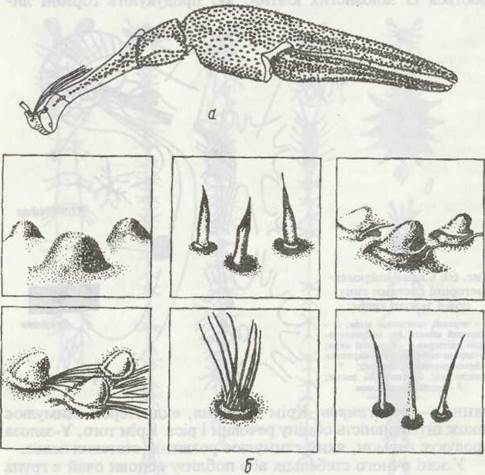
Рис. 61. Схема розміщення рецепторів на поверхні кінцівки річкового рака (а) та зовнішній вигляд кутикулярної частини рецепторів (б)
Ракоподібні, як і інші членистоногі, крім центральної, мають ще вегетативну нервову систему, яка тісно пов'язана з центральною. Вона іннервує внутрішні органи і впливає на діяльність травної, дихальної, видільної, кровоносної систем, а також на обмін речовин в організмі. Найкраще вивчено передній (стоматогастричний) відділ вегетативної нервової системи (див. рис. 58). У річкового рака він складається з глоткового, шлункового гангліїв та пари гангліїв, що лежать на навкологлоткових конективах. Від гангліїв відходять нерви до внутрішніх органів.
Органи чуття розвинені добре. Тверді ділянки кутикули ракоподібних, як і в інших членистоногих, нечутливі до подразнень, і чуття дотику та хімічне чуття приурочене до певних структур —сенсил (див. рис. 49). Кутикулярна частина сенсили має вигляд волосинки, щетинки, коротенького конуса або ямки. Дотичні сенсили розташовані здебільшого на антенулах, антенах та інших кінцівках (рис. 61). На антенулах у багатьох раків є органи хімічного чуття (смаку) у вигляді гребінця з особливих тонкостінних щетинок (рис. 62).
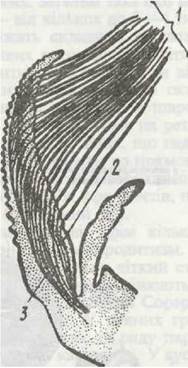
Рис. 62. Чутливі щетинки на антенулі краба Geryon affinis:
1 - гребінець чутливих щетинок; 2 - чутливі клітини; 3-нерв
Органи рівноваги відомі тільки в деяких вищих раків. У Decapoda в базальному членику антенул є особлива камера — статоцист, внутрішні стінки якої мають особливі волоскоподібні рецептори (рис. 63). Статоцист з'єднаний із навколишнім середовищем отвором, через який усередину потрапляють піщинки, що виконують роль статолітів. При линянні вони випадають, і рак відновлює запас статолітів, підбираючи їх кінцівками або занурюючи голову в пісок. У мізид статоцисти розташовані в останній парі черевних ніжок (уроподах).
Органи зору в ракоподібних бувають двох типів. У багатьох із них є непарне наупліальне око, розташоване між основами антенул, характерне як для личинок (наупліуса), так і для багатьох дорослих ракоподібних (гіллястовусі, копеподи тощо). Кожне наупліальне око (рис. 64, а) складається з трьох-чотирьох злитих утворів — бокалів. Бокал складається зі спеціальної прозорої лінзової клітини, що править за кришталик, та розташованого під ним шару з кількох світлочутливих (ретинальних) клітин. Знизу та з боків він оточений пігментними клітинами. У різних груп будова наупліальних очей істотно відрізняється за кількістю бокалів, клітинним складом; часто відсутня лінза. Ці очі належать до так званих інвертованих: нервові відростки відходять від ретинальних клітин з боку лінзи. Вважають, що наугогіальні очі здатні реагувати на ступінь освітленості, однак предметів не бачать.
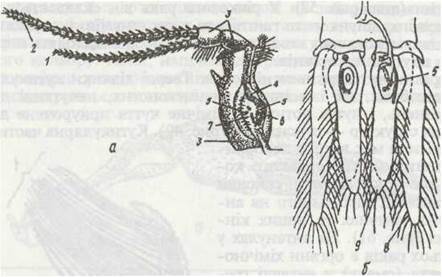
Рис. 63. Статоцисти:
а - в антенулі річкового рака; 6 - в уроподах мізид; І, 2 -джгути антенули; 3 - основні членики; 4 - отвір ямки статоциста; 5 - чутливі волоски на дні статоциста; б — піщинки; 7 - нерв статоциста; 8 - екзоподит; 9 — ендоподит
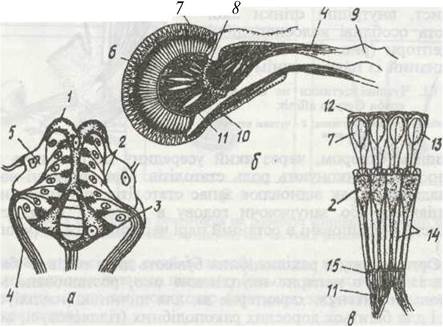
Рис. 64. Схема будови очей ракоподібних:
а - наупліальне око Ostmcoda, б - фасеткове око Branchipus; в - окрема його ділянка; 1 - бокал; 2 - ретинальні клітини; 3 - пігментні клітини; 4 - зоровий нерв; 5 - кришталик; 6 - шар ретинальних і пігментних клітин; 7 - кришталеві конуси; 8 - гангліозні скупчення нервових клітин; 9-мозок; 10 - м'яз очного стебельця; 11 -нервові волокна; 12 -прозора кутикула; 13 - оматидій; 14 - пігментний прошарок; 15 - базалвна мембрана
Більшість дорослих ракоподібних здатна розрізняти предмети за допомогою складних (фасеткових) очей, які характерні також і для інших членистоногих. Загалом таке око має багато окремих вічок (оматидіїв) — від кількох до 3—4 тис. (рис. 64, б, в). До оматидіїв належать складний світлоза- ломлювальний апарат, групи чутливих ретинальних клітин, від яких відходять нерви, та пігментних клітин. Нерви відходять від ретинальних клітин знизу (неінвертоване око). Кожен оматидій ізольований від інших пристінним шаром пігменту, який поглинає більшість променів. Тому на ретинальні клітини потрапляє лише частина променів, що падають на поверхню оматидія під кутом, близьким до прямого. Отже, кожен оматидій сприймає лише частину зображення. Суцільне зображення складається з багатьох фрагментів, немовби із часток мозаїчної картини (мозаїчний зір).
Більшість ракоподібних роздільностатеві, крім кількох сидячих форм, для яких характерний гермафродитизм. У роздільностатевих видів нерідко спостерігається чіткий статевий диморфізм. Так, деякі кінцівки самця видозмінюються в хапальні органи для утримання самиці (антенули Сорероda), або стають копулятивними органами. У певних груп самці значно менші, ніж самиці, в усоногах та ряду паразитичних рівноногих ракоподібних самці карликові. У вусоногих раків спостерігається гетерогонія — чергування партеногенетичних поколінь із двостатевими.
У небагатьох ракоподібних (Isopoda та ін.) гонади парні, кожна з них сполучається із зовнішнім середовищем за допомогою статевої протоки. Частіше гонади зливаються, однак статеві протоки та отвори залишаються парними (рис. 65). Від яєчника відходять яйцепроводи із залозистими стінками, які виділяють навколо яєць щільну шкаралупу. Часто у самиць є сім'яприймачі, куди потрапляє сперма. Це парні ампули, що відкриваються назовні поблизу отворів яйцепроводів спеціальними отворами. У сім’яприймачах сперма зберігається доти, доки з яйцепроводів не виходять зрілі яйця; в цей час відбувається запліднення.
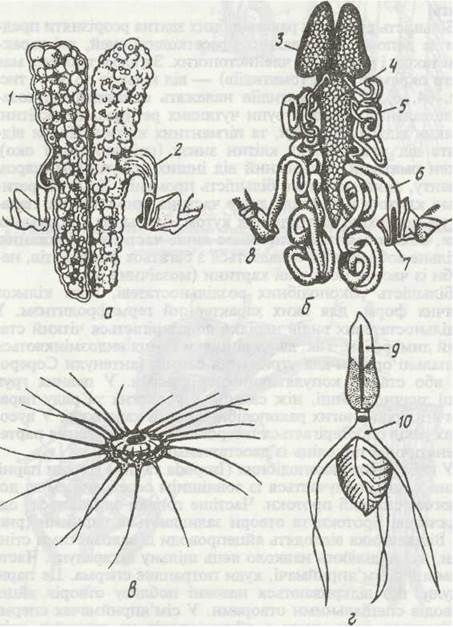
Рис. 65. Десятиногі:
а - жіноча, б - чоловіча статеві системи річкового рака; в — його сперматозоїд; г - сперматозоїд Galathea;
1 - яєчник; 2 - яйцепровід; 3 - парна частина сім'яника; 4 - непарна частина сім'яника; 5 -сім'япровід; 6 - сім'явиносний канал; 7-статевий отвір; 8 - протоподит задньої грудної ноги; 9 -хвостова капсула; 10 - шийка з трьома відростками
Стінки сім'япроводів, які відходять від сім'яників, також мають залозисті клітини, що виділяють оболонку сперматофорів; інколи сім'япроводи розширюються в сім'яні пухирці, де остаточно формуються сперматофори.
У більшості ракоподібних сперматозоїди типової форми, але можуть (наприклад, Ostracoda) досягати величезної довжини, до 5—6 мм (найдовші сперматозоїди, відомі у тварин). У багатьох вищих раків сперматозоїди мають досить химерну форму й зовсім нерухомі. Так, у річкового рака вони схожі на диск із променеподібними відростками, а у Galatheaскладаються з двох овальних мішків (один із них на кінці загострений), сполучених вузькою шийкою, від якої в один бік відходять три довгі відростки (рис. 65, в, г). Мішок із загостреним кінцем відповідає голівці звичайного сперматозоїда, а другий мішечок — хвостовій частині.
Плодючість ракоподібних дуже різна: річковий рак відкладає близько 200 яєць, а деякі краби — до двох мільйонів.
Яйця ракоподібних одягнені щільною оболонкою —хоріоном і, як правило, виношуються на ніжках самиці або в особливих виводкових камерах. Іноді вони склеюються у яйцеві мішки, прикріплені до тіла.
Яйця найчастіше багаті на жовток, проте бувають і з незначною його кількістю (деякі Copepoda та ін. ).
Яйця з невеликою кількістю жовтка проходять повне, нерівномірне, детерміноване дробіння, яке нагадує таке у кільчастих червів. У яйцях з великою кількістю жовтка дробіння інше: воно стає неповним і поверхневим (рис. 66), причому спочатку багаторазово ділиться ядро яйцеклітини. Ядра, що утворились, мігрують на периферію яйця, й лише після цього навколо кожного з них відокремлюється ділянка цитоплазми. Утворюється один шар клітин, що оточує центральну масу жовтка (стадія, що відповідає бластулі, порожнина якої заповнена жовтком).
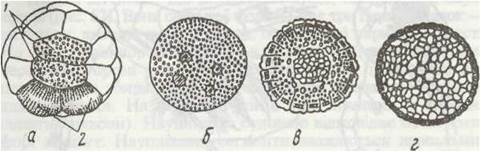
Рис. 66. Дробіння яйця ракоподібних:
а - повне детерміноване дробіння Euphausia; б-г три стадії поверхневого дробіння краба Diomia;
(г - стадія, що відповідає бластулі); 1 - ентодерма; 2 - первинні мезодермальні клітини
Потім на одному полюсі бластули виникає багатошарова пластинка — зародкова смужка, з якої розвивається зародок. Вона сегментується, на її передньому кінці закладаються парні головні лопаті (зачатки протоцеребрума), складні очі й одне непарне просте (наугогіальне) око. На черевній стороні зародка формуються три пари кінцівок (антенули, антени і мандибули). Мезодерма зародка має вигляд двох мезодермальних смужок, які інколи сегментуються, як у кільчаків, але згодом целомічні мішки руйнуються, а їхні стінки утворюють мезодермальні органи (м’язи, серце з судинами, органи виділення). Целомічні порожнини зливаються із залишками первинної порожнини тіла, утворюючи змішану порожнину тіла — міксоцель. Іноді сегментація мезодерми не має чіткості, а справжній целом взагалі не формується.
Більшість ракоподібних розвивається з метаморфозом. У різних груп він відбувається по-різному.
Перша личинка, що виходить з яйця, називається науп- ліусом (рис. 67). Вона овальної форми і має три пари кінцівок — антенули, антени та мандибули; останні дві пари двогіллясті. На відміну від дорослих форм, у наупліуса антени лежать на черевній стороні тіла, по боках від рота, і призначені для плавання та збирання іжі, а мандибули спочатку — тільки для плавання. На задньому кінці тіла личинки є анальна лопать (тельсон). Наупліус за будовою відповідає метатрохофорі поліхет. Наупліальні сегменти вважаються ларвальними, всі інші, що виникають пізніше, постларвальними. Щодо кількості ларвальних сегментів у ракоподібних єдиної думки немає.
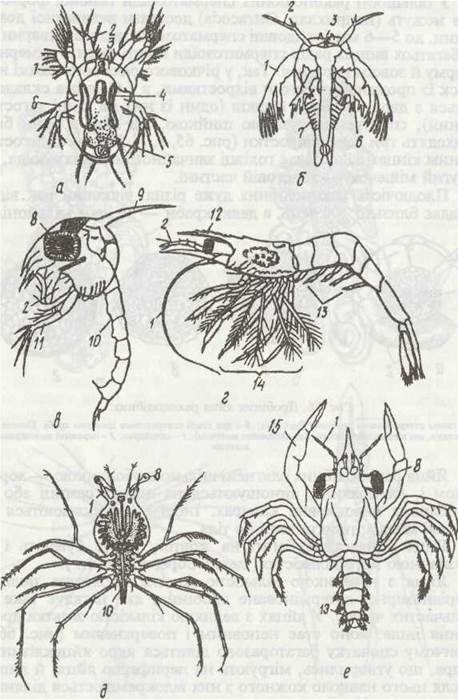
Рис. 67. Личинкові стадії ракоподібних:
а - наупліус Cyclops; б - метанаупліус Арm; в - зоеа краба Maja; г - мізидна личинка Pandalus; д - філосома лангуста Palinunis; е - мегалопа краба Portunus; 1 - антени; 2- антенули; 3 - наупліальне око; 4 - антенальиа залоза; 5 - кишечник; 6 - макдибула; 7 - грудні сегменти; 8 - фасеткове око; 9 - спинний шип; 10 - черевні сегменти; 11 - ногощелепа; 12 - рострум; 13 - черевні ноги; 14 - грудні ноги; 15 - клішня
Наупліус має кишечник, головний мозок, два черевні ганглії, непарне наупліальне око й одну пару видільних органів, найчастіше це антенальні залози. На задньому кінці тіла, між мандибулярним сегментом і тельсоном, розташована зона росту. Тут утворюються наступні сегменти тіла.
Наупліуси ведуть планктонне життя й розносяться (особливо морськими течіями) на далеку відстань.
Наупліус росте, в зоні росту утворюються нові сегменти із зачатками кінцівок: спочатку сегменти з першою і другою парою максил, потім — передні грудні. Личинка, в якої починають формуватися постларвальні сегменти, зветься метанаупліусом.
Під час постембріонального розвитку відбуваються линяння. Після кожного з них з'являються нові сегменти і закладаються нові кінцівки, тобто розвиток відбувається шляхом анаморфозу. При цьому можуть змінюватися будова та функції кінцівок. Наприклад, усі три пари кінцівок наупліуса, як уже згадувалося, призначені для плавання; пізніше ж антенули і антени стають органами чуття, а мандибули — ротовими придатками. Поступово кінцівки набувають остаточного вигляду, і метаморфоз закінчується.
Постембріональний розвиток у класі Malacostraca має свої особливості. У них скорочується й стабілізується кількість линянь, утворюються додаткові личинкові стадії. Часто спостерігається ембріонізація розвитку, в результаті якої ранні личинкові стадії (наупліус та метанаулліус) проходять у яйці, а назовні виходить більш пізня личинка (найчастіше зоеа). Зоеа — це личинка, що вже має всі сегменти, але середні сегменти ще повністю не сформовані й не мають кінцівок. Зоеа має розвинені ротові кінцівки й ногощелепи, а також зачатки грудних ніжок і сформоване черевце, на якому є лише остання пара кінцівок. На передній частині тіла є фасеткові очі. Зоеа линяє й переходить у так звану мізидну стадію: у неї вже є повністю сформовані двогіллясті грудні ноги (як в представників ряду Mysidacea, звідки й назва личинки) й зачатки черевних кінцівок (рис. 67, в, г).
У різних представників десятиногих раків мізидна стадія має різну будову і відповідно різні назви: у крабів-мегалопа, що зовні нагадує дорослу тварину, але має витягнуте черевце, у лангустів — фіпосома, яка має прозоре листкоподібне тіло, довгі двогіллясті грудні кінцівки й дуже коротке почленоване черевце тощо (рис. 67, д, е). Мізидна стадія після линяння перетворюється на цілком сформовану тварину.
Деякі вищі раки, наприклад креветки з родини Penaeidae, проходять усі личинкові стадії: наупліус, метанаулліус та наступну мізидну стадію. У ряді груп метаморфоз відбувається дуже своєрідно, про що буде сказано при розгляді цих груп.
У більшості видів ряду Decapoda, а також у деяких інших таксонах спостерігається повна ембріонізація розвитку, в результаті чого з яйця виходить мініатюрна копія дорослої тварини з усіма сегментами й сформованими кінцівками, тобто вони розвиваються шляхом епіморфозу.
Ріст раків супроводжується періодичними линяннями, в яких беруть участь не тільки покриви, а й внутрішні органи, нервова й ендокринна системи. Найкраще цей процес вивчено у річкового рака. У передлиняльний період стара кутикула відшаровується від гіподерми, її внутрішні шари розсисаються, а речовини, що звільнилися при цьому, відкладаються у внутрішніх органах. У бічних стінках шлунку утворюються округлі тверді бляшки, що звуться гастролітами, або «жорновками». Вони складаються в основному із солей кальцію, які звільняються при руйнуванні старої кутикули, а потім використовуються при затвердінні нової кутикули. У тканинах і гемолімфі нагромаджуються речовини, необхідні для побудови нової кутикули: у гіподермі — глікоген, у печінці — мінеральні речовини. Паралельно цьому клітинами гіподерми виділяється нова кутикула. Речовини, необхідні для побудови кутикули (вуглеводи, ліпіди тощо), надходять із гемолімфи і тканин. У цей період в організмі підвищується інтенсивність процесів обміну речовин, про що свідчить посилене споживання кисню.
Власне процес линяння полягає в тому, що стара кутикула лопається в певному місці, і через цю щілину тварина звільняється від старої кутикули, яка залишається у вигляді порожнього чохла — екзувія, котрий цілком зберігає форму тіла рака та його придатків. Нова кутикула спочатку м'яка, еластична і не має характерного для даного виду забарвлення. У цей період збільшуються розміри тіла за рахунок нагромадження в тканинах води та їх набрякання. Через деякий час (протягом кількох годин) кутикула твердіє (склеротизація), і в ній нагромаджуються мінеральні солі, які звільняються при розчиненні гастролітів, конкрецій у клітинах печінки, а також надходять з їжею.
До підтипу Зябродишні, або Ракоподібні, за сучасною системою належать 6 класів, шо включають 12 підкласів та 38 рядів. Далі ми розглянемо найголовніші з них.