ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 3 - Г.Й. Щербак - 1997
ТИП МОЛЮСКИ, АБО М’ЯКУНИ (MOLLUSCA)
КЛАС ЧЕРЕВОНОГІ (GASTROPODA)
Черевоногі — найбагатший за кількістю видів клас молюсків: їх близько 90 тис. В Україні відомо понад 500 видів. Черевоногі мешкають у різних біотопах Світового океану — від берегової зони до глибин більше 10 тис. м; у прісних водоймах та на суходолі; від полярних широт до тропіків; від рівнин до гірських вершин (понад 5 тис. м над рівнем моря). Дуже невелика кількість видів веде паразитичний спосіб життя.
Розміри черевоногих варіюють від 2—3 мм до кількох десятків сантиметрів. Найкрупніші з них: Hemifusus proboscidiferus з черепашкою завдовжки 60 см; морський заєць — Aplysia depilans — розміром 40 см; деякі африканські види наземних слимаків роду Achatina завдовжки до 25 см; червоподібний ендопаразит голотурій Parenteroxenos dogeli завдовжки 128 см.
Характерною рисою класу Gastropoda є асиметричність будови, яка виражається у формі черепашки, редукції органів мантійного комплексу однієї сторони (здебільшого правої) та посиленим розвитком таких самих органів іншої сторони (здебільшого лівої).
Тіло черевоногих складається з голови, ноги та тулуба, який утворює виріст — нутрощевий мішок, вкритий суцільною ковпачкоподібною або спірально закрученою черепашкою (рис. 56).
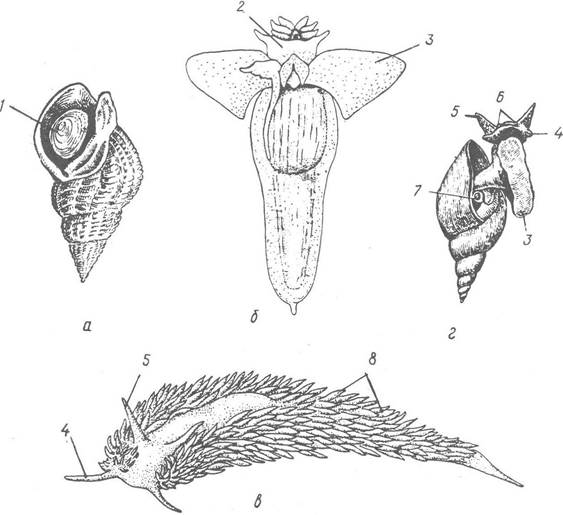
Рис. 56. Черевоногі молюски:
а — Buccmum undatum (підклас Prosobiauchia); б — морський ангел (Clione limacina, підклас Opisthobranchia, ряд Pteropoda); в — Aeolidia papillosa (підклас Opisthobranchia, ряд Nudibranchia); г — звичайний ставковик (Limnaea stagnalis, підклас Pulmonata, ряд Basommatophora); 1 — кришечка; 2 — голова; 3 — нога; 4 — ротова лопать; 5 — щупальце; 6 — очі; 7 — дихальний отвір; 8 — адаптивні зябра
Голова чітко відокремлена від тіла, на ній розташовані рот, одна або дві пари щупалець та пара очей (рис. 57, а). У деяких форм голова витягується І утворює так зване рило, а в деяких хижих та паразитичних Prosobranchia перетворюється на мускулястий хоботок, який може вгортатись або викидатись назовні при захопленні здобичі.
Нога добре розвинена і здебільшого має підошву, пристосовану для повзання. У плаваючих форм бічні краї підошви розростаються, утворюючи широкі лопаті (наприклад, у Арlysia, ряд Tectibranchia) або плавці, як у Сlionе limacina (ряд Pteropoda), за допомогою яких ці тварини плавають (див. рис. 56). У деяких паразитичних форм нога редукується. Над ногою міститься мішкоподібний тулуб, або нутрощевий мішок, вкритий черепашкою.
Тулуб, або нутрощевий мішок, у нижчих черевоногих (наприклад, деяких передньозябрових), голозябрових підкласу задньозяброві та деяких легеневих симетричний і нечітко відділений від ноги. У більшості ж черевоногих тулуб, навпаки, видається над ногою у вигляді великого, більш-менш закрученого спірально мішка.
Черепашка суцільна, ковпачкоподібна або здебільшого закручена спіраллю за годинниковою стрілкою, тобто праворуч, якщо дивитись на неї із загостреного кінця (дексіотропні черепашки), рідше трапляються лівозакручені (леотропні) черепашки. Закрутки черепашки, щільно прилягаючи один до одного, можуть зростатися своїми внутрішніми стінками, утворюючи суцільний стовпчик, який називається колонкою (рис. 57, б), а іноді можуть відставати один від одного. Тоді по осі черепашки утворюється щілина — так званий пупок.
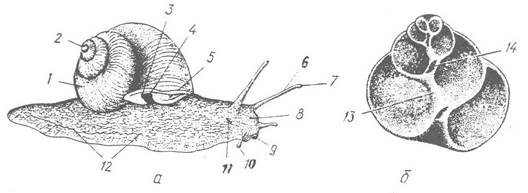
Рис. 57. Виноградний слимак (Helix pomatia):
а — зовнішній вигляд з правої сторони; б — поздовжній розпил черепашки; 1 — черепашка; 2 — верхівка черепашки; 3, 4 — анальний та дихальний отвори; 5 — край мантії; 6 — очне щупальце; 7 — око; 8 — голова; 9 — рот; 10—губне щупальце; 11—статевий отвір; 12 — нога; 13, 14 — стовпчик та його порожнина
На одному кінці (верхівці) черепашка сліпо замкнена, а на протилежному є отвір (вустя), через який висуваються назовні голова й нога тварини. Лише у виняткових випадках закрутки спіралі черепашки лежать в одній площині (планоспіраль), як, наприклад, у прісноводних котушок (рід Ріа- norbis); у більшості ж спіраль конічна (турбоспіраль). Висота турбоспіралі в різних видів різна, діаметр закрутків збільшується від верхівки до вустя. В інволютних черепашок кожний новий, більший, закруток охоплює всі попередні, робля чи їх непомітними (наприклад, Сургаеа). В еволютних черепашок останній закруток лише прилягає до попередніх, не закриваючи їх (Helix).
Черепашка може мати й вигляд конічного ковпачка, або блюдечка, як, наприклад, у морських блюдечок (Patella та Астеа, див. далі) та річкових чашечок (Ancylus). Проте їхні личинки спочатку мають більш-менш закручену черепашку, яка лише пізніше набуває вигляду ковпачка.
У більшості випадків черепашка настільки велика, що в неї ховається все тіло молюска. У деяких форм, переважно з підкласу Prosobranchia, на спинній стороні ноги утворюється вапнякова або рогова пластинка — кришечка, якою при втягуванні тіла в черепашку замикається вустя (див. рис. 56, а).
У черевоногих часто спостерігається редукція черепашки, а іноді й повне її зникнення. В одних випадках черепашка зменшується в розмірах і прикривається мантією або бічними ділянками ноги і стає внутрішньою (Aplysia, слизун Limax), в інших —розпадається на окремі вапнякові тільця, що лежать у покривах спинної сторони (слизун Arion — ряд Nudibranchia, див. рис. 56). Інколи навіть сліди черепашки зникають (Pteropoda). Редукція черепашки спостерігається в плаваючих і наземних форм, що полегшує їх тіло, а також у паразитичних черевоногих.
Черепашка черевоногих, як і інших м’якунів, складається з тонкого органічного зовнішнього шару (периостракума), під яким залягає порцеляноподібний шар (остракум), у деяких черевоногих (Haliotis, Turbo та ін.), є ще внутрішній перламутровий шар (гіпостракум).
На тулубі утворюється складка покривів, мантія, під якою міститься мантійна порожнина з розташованим у ній мантійним комплексом органів (зябра, анальний отвір, видільний та статевий отвори). Як мантія, так і мантійна порожнина звичайно розвинені на передній і правій сторонах тулуба. Мантія виділяє черепашку. Ріст черепашки протягом життя тварини відбувається по її потовщеному вільному краю, який містить безліч залозистих клітин.
Шкіра черевоногих складається з одношарового покривного епітелію, або епідермісу, підстеленого базальною мембраною, і сполучної тканини (кутиса), яка лежить під ним. Епітеліальні клітини зовні виділяють тонку кутикулу, і тільки на підошві ноги й навколо дихального отвору в Pulmonata кутикули немає; у цих ділянках епітелій війчастий (рис. 58).
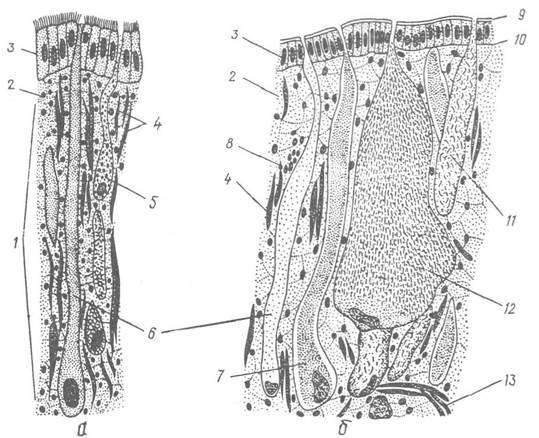
Рис. 58. Зріз через шкіру підошви ноги (а) та краю мантії (б) Helix pomatia:
1 — купе; 2 — сполучна тканина; 3 — покривний епітелій; 4 — дорзовентральні м’язові волокна; 5, 6, 7 — пігментна, слизова та білкова залози; 8 — поздовжні м’язові волокна; 9 - кутикула; 10 — базальна мембрана; 11, 12 — слизова мантійна та вапнякова залози; 13 — поперечні м’язові волокна
Шкіра багата на залозисті клітини, які занурені під епідерміс у сполучну тканину. їх протоки відкриваються назовні між клітинами епідермісу. Залози бувають слизовими, білковими та пігментними. Слизових залоз особливо багато на підошві ноги й по краю мантії, де вони досягають величезних розмірів. Слиз змащує тіло м’якуна, зволожуючи і захищаючи його, а на нозі він сприяє повзанню. Шкіра молюска завжди зволожена слизом. Білкові залози містяться в шкірі тулуба та по краю мантії. Особливою різновидністю білкових залоз є вапнякові залози, які досягають великих розмірів на краю мантії. їх секрет використовується для побудови епіфрагми — плівки, якою затягується вустя черепашки при несприятливих умовах. Пігментні залози містяться в шкірі мантії та інших частин тіла, особливо щупалець.
Сполучна тканина складається із зірчастих клітин паренхіми, міжклітинної речовини та ніжних волокнистих тяжів, що переплітаються в різних напрямах з мускульними пучками. Особливо багата на мускулатуру сполучна тканина ноги. Тут містяться численні лакуни.
Крім уже зазначених м’язів шкіри, у черевоногих добре розвинена мускулатура ноги, яка складається з поздовжніх, кільцевих, діагональних та дорзовентральних м’язів. Є також спеціалізовані м’язи, серед яких найбільше розвинений колу- молярний м'яз, що втягує тварину в черепашку. Верхній кінець його прикріплюється до стовпчика (columella) черепашки, а нижній розпадається на окремі пучки, які втягують у черепашку голову, щупальця та ногу молюска (рис. 59). Якщо черепашка редукується, цей м’яз також редукується, або в плаваючих форм виконує локомоторну функцію. У черевоногих є також складно диференційована мускулатура ротових органів та копулятивного апарату.
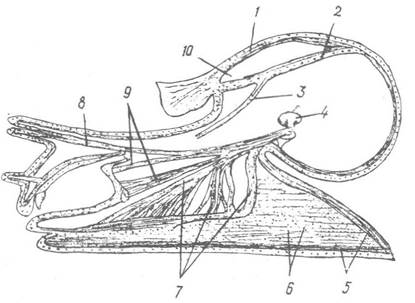
Рис. 59. Схема будови мускулатури Helix pomatia:
1 — поздовжні та 2 — кільцеві м’язи шкіри; 3 — ретрактор копулятивного органа; 4 — стовпчик; 5 — поздовжні та 5 — косі м’язи ноги; 7 — 9 — ретрактори ноги, задніх щупалець та глотки відповідно; 10 — легенева порожнина
Целом у черевоногих невеликий, складається з двох самостійних утворень — порожнини гонади та перикардія. Порожнина тіла, в якій міститься передня частина травної та статева системи — це великий венозний синус, який є ділянкою первинної порожнини тіла (схізоцелем). Паренхіма, на відміну від двостулкових, у черевоногих розвинена слабо. Вона утворює сполучнотканинний шар покривів та оточує печінку й нирки.
Більшість черевоногих живляться перифітоном, який зішкрібають із підводних предметів, а наземні — м’якою тканиною з живого або гниючого листя. Є серед них і хижаки.
Рот міститься на нижній стороні переднього кінця голови і часто оточений шкірними складками, або губами. Він веде в ротову порожнину, яка переходить у мускулясту глотку (рис. 60). На межі глотки й ротової порожнини дорзально лежать одна або дві щелепи. У виноградного слимака щелепа має вигляд вигнутої півмісяцем пластинки з поперечними
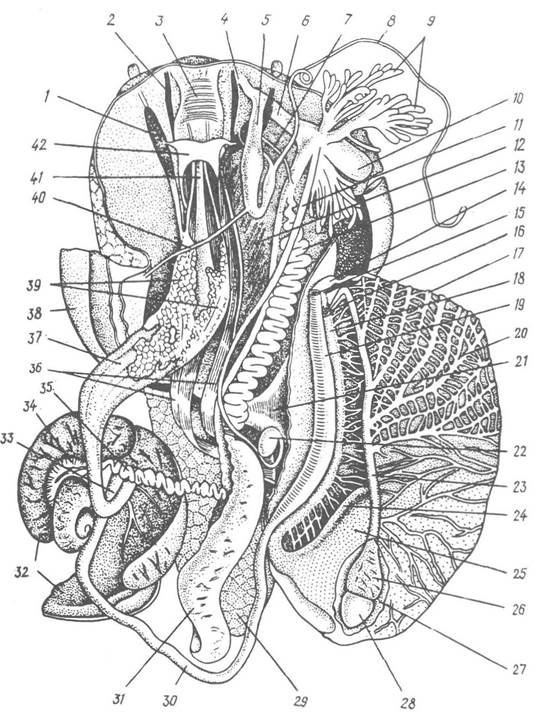
Рис. 60. Внутрішня будова Helix pomatia:
1,2 — очне та губне щупальця, втягнуті всередину; 3 — глотка; 4 — статева клоака; 5 — мішок копулятивного органа; б — піхва; 7 — сім’япровід; 8 — бич; 9 — пальцеподібні залози; 10 — мішок любовної стріли; 11 — канал сім’яприймача; 12 — ретрактори ноги; 13 — сім’яяйцепровід; 14 — край дихального отвору; 15 — анус; 16 — видільний отвір; 17 — приносна легенева судина; 18 — вторинний сечовід; 19 — пряма кишка; 20 — виносна легенева судина; 21 — колумелярний м’яз; 22 — сім’яприймач; 23 — легенева вена; 24 — первинний сечовід; 25 — нирка; 26 — передсердя; 27 — перикардій; 28 — шлуночок; 29 — задній кінець ноги; 30 — тонка кишка; 31 — білкова залоза; 32 — печінка; 33 — шлунок; 34 — гермафродитна залоза; 35 — гермафродитна протока; 36 — ретрактори голови, глотки та щупалець; 37 — воло; 38 — мантія; 39 — слинні залози; 40 — ретрактор копулятивного органа; 41 — стравохід; 42 — церебральний ганглій
реберцями, яка вдається в ротову порожнину (рис. 61). Щелепи рогові, іноді просякнуті вуглекислим кальцієм. Стінки глотки тверді й мускулясті з твердою кутикулярною вистилкою всередині. Ззаду глотка сполучається з радулярною піхвою. Із дна глотки вип’ячується язик, на поверхні якого є тертка (радула). Радула складається з основної кутикулярної пластинки, поверхня якої вкрита численними поперечними рядами рогових зубців, спрямованих вістрями назад (рис. 62). Ближче до переднього кінця язика зубці тверді й гострі, а на задньому кінці, біля основи язика — ніжні й м’які. Зубці радули на передньому краї стираються, а на задньому весь час відновлюються. Як і в хітонів, у черевоногих до складу зубців радули входить залізо.
Рис. 61. Щелепа Helix pomatia
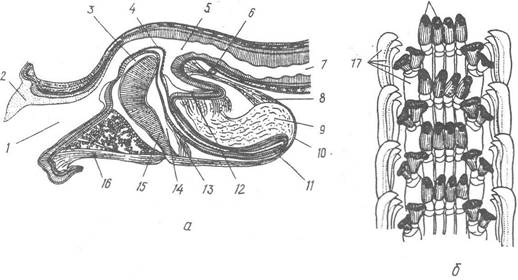
Рис. 62. Будова глотки та радули черевоногих:
а — медіальний зріз глотки Helix pomatia; б — частина радули Patella pontica; 1 — ротова порожнина; 2 — щелепа; 3 — язик; 4 — радула; 5 — порожнина глотки; 6 — букальна комісура; 7 — стравохід; 8, 9 — м’яз, що підтримує радулярну піхву та сполучна тканина піхви; 10 — радулярна піхва; 11 — епітелій, що утворює радулу; 12 — складка епітелію глотки; 13 — внутрішній глотковий м’яз; 14 — кровоносні лакуни язика; 15 — радулярний хрящ; 16 — мускульна стінка глотки; 17 — бічні зуби; 18 — серединні зуби
У глотку відкриваються протоки однієї пари слинних залоз, а в деяких слимаків — протоки інших залоз, наприклад отруйних, або залоз, які виділяють кислоту. Секрет слинних залоз містить слиз, який зволожує і змащує харчову масу, та ферменти, що розщеплюють крохмаль, а в хижих форм — білки.
Глотка переходить у стравохід, який у деяких м’якунів утворює розширення —воло. Усі зазначені органи належать до ектодермальної передньої кишки.
Ентодермальна середня кишка складається з мішкоподібного шлунка та довгої тонкої кишки. Травна залоза, або печінка, складається з численних часток, протоки яких з’єднуються і впадають у шлунок (див. рис. 60).
Шлунок молюсків має різну будову залежно від характеру живлення. У мікрофагів, тобто тих, що живляться дрібнодисперсною їжею (мікроорганізмами, детритом), шлунок має найскладнішу будову. Наприклад, у Fissurella (Prosobranchia) у шлунку розрізняють три функціональні зони: сортувальне поле, кутикулярний щит та мішок протостиля. Сортувальне поле вкрите війками і утворює складну систему складок і борозен. Харчові частинки ще в стравоході огортаються слизом і в шлунок потрапляють у складі слизового шнура, від якого там відлипають. Більші частинки, які потрапляють на сортувальне поле, заганяються в передкишкову борозну шлунка, а звідти — у кишку. Дрібні частинки залишаються в завислому стані в шлунку. Під дією травних ферментів слини вони частково перетравлюються, а далі потрапляють у печінку, де фагоцитуються клітинами її епітелію; тут відбувається внутрішньоклітинне травлення. На сортувальному полі, крім того, у просвіт шлунка проникає безліч амебоцитів гемолімфи, які також фагоцитують дрібні харчові частинки. Кутикулярний щит покриває одну із стінок шлунка. Мішок протостиля лежить у задній частині шлунка біля початку тонкої кишки. У ньому міститься паличка з ущільненого слизу (протостиль), обліплена великими частинками їжі та неперетравленими рештками, що повертаються з печінки. Протостиль обертається завдяки роботі війок свого мішка і втягує в шлунок зі стравоходу з’єднаний з ним слизовий шнур з їжею.
У деяких мікрофагів підкласу Prosobranchia протостиль перетворився на щільний, прозорий кришталевий стовпчик, як і в двостулкових (див. с. 41), до якого прилипають харчові частинки. Кінець його під впливом лужного середовища шлунка поступово розчиняється і звільнює ферменти, які розщеплюють вуглеводи їжі та клітинні оболонки водоростей.
Деякі з Prosobranchia (в основному хижі), а також більшість Pulmonata та Opisthobranchia мають простіший за будовою шлунок, у стінках якого залягають м’язи; слизово-війчастий механізм руху їжі по кишечнику змінився в них на перистальтику мускулатури стінок кишкового тракту. У деяких з них (наприклад, у ставковика Limnaea) ще залишаються рудименти сортувального поля та кришталевого стовпчика. У виноградного слимака подрібнена та змішана з секретом слинних залоз їжа надходить до вола, де піддається діїсекрету печінки, який розщеплює крохмаль та клітковину і омилює жири. Далі харчова маса проходить у шлунок і звідти по печінкових протоках — у печінку. Розчинені поживні речовини всмоктуються печінкою, а білкові частинки фагоцитуються клітинами печінки, піддаючись внутрішньоклітинному травленню.
У м’ясоїдних форм (Muricidae, Buccinidae та ін.) протеолітичні ферменти виділяються слинними залозами, травлення проходить у волі та порожнині шлунка. Отже, у черевоногих є всі переходи від внутрішньоклітинного травлення до порожнинного.
Печінка закладається у вигляді парного вип’ячування кишечника, але в дорослих черевоногих у зв’язку з їх асиметрією вона непарна — одна її половина (права) недорозвинена.
Отже, печінка черевоногих виконує складні функції: крім фагоцитозу дрібних частинок їжі, у хижих форм вона виділяє ферменти, які надходять у шлунок і навіть у воло; її епітелій всмоктує продукти травлення; вона є місцем відкладання та накопичення поживних та деяких інших речовин (глікоген, жири, фосфорнокислий кальцій). Жири, крім того, відкладаються в сполучній тканині, що оточує печінку.
Своєрідних змін зазнає печінка в голозябрових, де вона розбивається на систему залозистих каналів, гілочки яких заходять у щупальцеподібні вирости, які вкривають спину — вторинні зябра, і можуть навіть відкриватися назовні (рис. 63). У таких щупальцеподібних відростках містяться жалкі капсули (наприклад, у представників родини Aeolididae). Спеціальні дослідження показали, що жалкі капсули гідроїдів, якими живиться молюск, не перетравлюються, а надходять до печінкових виростів, зберігаючи свої захисні функції.
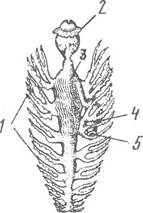
Рис. 63. Кишечник Aeolis (ряд Nudibranchia):
1 — розгалужена печінка; 2 — глотка; 3 — шлунок; 4 — задня кишка; 5 — анус
Від шлунка відходить довга тонка кишка, яка в багатьох черевоногих має внутрішній поздовжній виріст — тифлозоль. У тонку кишку із шлунка та печінки надходять неперетравлені рештки їжі. Травлення та всмоктування тут не відбуваються. Основна функція тонкої кишки — формування екскрементів та огортання їх слизом, що дуже важливо для слимаків, в яких анус відкривається в мантійну порожнину де містяться зябра, або поруч із дихальним отвором (у легеневих). Кишка утворює одну або кілька петель, а потім повертає вперед і переходить в ектодермальну задню кишку; остання відкривається порошицею на передньому кінці тулуба над головою або на правому боці тіла. У деяких нижчих черевоногих задня кишка проходить крізь шлуночок серця.
Видільна система складається з пари нирок, що є видозміненими целомодуктами. У примітивних форм зберігаються права та ліва нирки, а в більшості залишається лише ліва. Нирка має вигляд великого мішка (див. рис. 60), один кінець якого відкривається в перикардій (ділянка целома), від її другого кінця відходить довгий сечовід, який відкривається видільним отвором у мантійну порожнину. Внутрішня порожнина нирки вкрита складками, які збільшують її поверхню, а стінки обплетені численними кровоносними судинами.
Кровоносна система черевоногих має складну будову. Серце в різних систематичних групах відрізняється будовою та положенням, що пов’язано з розташуванням та будовою органів дихання. Найпримітивніші форми підкласу Prosobranchia з двома симетричними зябрами мають симетричне серце, яке складається з шлуночка та двох передсердь і міститься по серединній лінії тіла на передньому кінці за головою; через шлуночок проходить задня кишка. У форм з більш-менш редукованою правою зяброю редукується й праве передсердя. У деяких видів праве передсердя зменшене і замкнене, що пов’язано з редукцією правої зябри та зябрової вени, яка впадає в це передсердя. У всіх інших слимаків (вищі представники підкласу Opisthobranchia та Pulmonata) зберігається лише ліве передсердя, тоді як праве повністю зникає. Положення передсердя, що залишилося, залежить від розташування зябри або легені (рис. 64). У передньозябрових та легеневих воно залишається попереду шлуночка, а в задньозябрових опиняється позаду нього, оскільки зябра зміщується далеко назад. Здебільшого серце лежить над задньою кишкою, у деяких примітивних Prosobranchia (Rhipidoglossa) шлуночок пронизує задня кишка. Серце оточене перикардієм (ділянка целома).
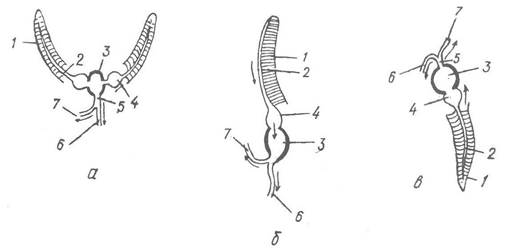
Рис. 64. Схема розташування ктенідій, серця та аорти в різних черевоногих:
а, б — види підкласу Prosobranchia з двома та одним ктенідіями відповідно; в — види підкласу Opisthobranchia; 1 — ктенідій; 2 — виносна зяброва судина; 3 — шлуночок серця; 4 — передсердя; 5 — аорта; 6 — нутрощева аорта; 7 — головна аорта
Від серця бере початок одна передня аорта, яка потім поділяється на головну аорту, що несе кров до переднього кінця тіла, та нутрощеву аорту, від якої відходять багато гілок (артерій) до різних органів (кишечника, печінки, статевої залози тощо). Вищі черевоногі мають дуже розгалужену артеріальну систему. Дрібні гілочки артерій переходять у дрібні капіляри, які розгалужуються в усіх органах. З них кров потрапляє в лакуни (щілини в паренхімі), особливо розвинені в нозі та по краю мантії, а потім збирається в більш-менш великі венозні синуси, що оточують кишечник, печінку та статеву залозу. Синуси є ділянками первинної порожнини тіла (схізоцеля), відмежованими сполучнотканинними мембранами, і з’єднані між собою через пори в цих мембранах. Найбільший з венозних синусів — це тулубний, в якому лежить передній відділ травної системи; крім того, є великий вісцеральний синус, ниркові та ректальний синуси.
Із синусів венозна кров надходить у вени, проходячи при цьому крізь нирку, де звільнюється від екскретів, а також через зябру, де насичується киснем, і врешті-решт потрапляє до передсердя, а звідти — у шлуночок. Отже, серце наповнюється переважно артеріальною кров’ю, лише незначна її частина надходить у передсердя, минаючи органи дихання.
У Pulmonata кров із великих синусів надходить у кільцевий синус, який оточує легеню, а звідти — у сітку судин, які пронизують стінку легені. Окиснена кров збирається в легеневу вену, яка впадає у передсердя.
Кров здебільшого безбарвна, містить пігмент гемоціанін, до складу якого входить мідь, через що на повітрі кров синіє. У деяких видів, наприклад роду Planorbis, кров містить червоний гемоглобін. У крові є амебоїдні клітини.
Органи дихання черевоногих надзвичайно різноманітні. Більшість черевоногих живе у воді й дихає зябрами. Первинними органами дихання є одна пара ктенідіїв, які лежать у мантійній порожнині. Характерною особливістю ктенідіїв є те, що біля основи кожного з них розташований орган хімічного чуття — осфрадій. Ктенідій має вигляд видовженого двопірчастого придатка, який складається з осьового стрижня з двома рядами зябрових пелюсток.
У найбільш примітивних Prosobranchia (наприклад, Puncturellа з родини Fissurellidae), два однаково розвинених ктенідія лежать у мантійній порожнині симетрично на передньому кінці тіла, над головою, і кінцями спрямовані вперед (рис. 65). У деяких Prosobranchia (Haliotis) правий ктенідій менший, ніж лівий, і, крім того, ктенідій по всій довжині приростає до стінки мантійної порожнини. В інших Gastropoda зберігається лише один лівий ктенідій, і він теж може видозмінюватись або редукуватись. У багатьох молюсків з двопірчастого він стає однопірчастим, приростаючи одним краєм до стінки мантії. Задньозяброві (Opisthobranchia) мають лише один ктенідій, зміщений назад по правій стороні тіла і обернений кінцем назад, а не вперед, як у Prosobranchia (див. рис. 64, а, б).
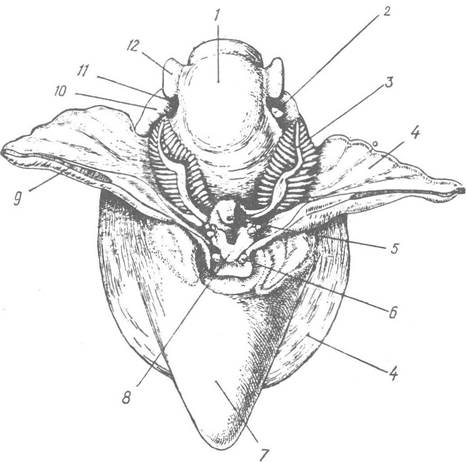
Рис. 65. Ктенідії в мантійній порожнині Punctuiella noachina (підклас Ргоsobranchia, черепашку видалено, мантійну порожнину розтято). Вигляд зі спинної сторони: 1 — голова; 2 — копулятивний орган; 3 — ктенідій; 4 — мантія; 5 — анус; 6 -— мантійне щупальце; 7 — нутрощевий мішок; 8 — сифональна лопать мантії; 9 — лінія, по якій розрізано мантію; 10 — передній край ноги; 11 — око; 12 — головне щупальце
У кожному підкласі Gastropoda є форми, в яких справжні зябра зникли і вторинно замінились іншими органами дихання. Наприклад, у чорноморських морських блюдечок (рід Patella підкласу Prosobranchia) по всьому краю мантії розвиваються численні складочки, які фізіологічно відповідають ктенідіям, а морфологічно є новоутвореннями, що називаються адаптивними зябрами (рис. 66). У голозябрових (ряд Nudibrancliia підкласу Opisthobranchia) адаптивні зябра також розташовані по краю мантії або утворюють віночок пірчастих виростів навколо ануса. У деяких з них (родина Aeolididae) зябрами є щупальцеподібні вирости спинної сторони тіла (див. рис. 56, в), про які вже згадувалося. Деякі Opisthobranchia зовсім не мають органів дихання і дихають всією поверхнею тіла (наприклад, Сііопе з ряду Pteropoda).
Наземні молюски підкласу легеневих (Pulmonata) перейшли до повітряного дихання, їхня мантійна порожнина перетворилася на легеню, заповнену повітрям. Край мантії в них зростається майже по всій довжині зі стінкою тіла, залишаючи лише невеличкий дихальний отвір, а на внутрішній стінці легені розвивається сітка численних кровоносних судин (рис. 67), через які відбувається газообмін. Деякі Pulmonata повернулися до життя в прісних водоймах, але в переважної більшості з них збереглося легеневе дихання. Відомі випадки, коли молюски мають подвійне дихання, наприклад прісноводна котушка (Planorbis corneus), в якої поряд із легенею є адаптивна зябра.
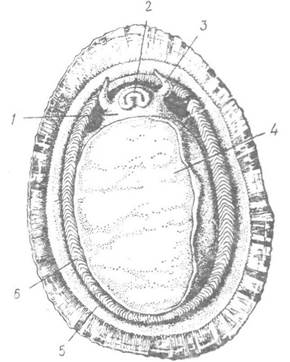
Рис. 66. Patella pontica (підклас Prosobranchia) з черевної сторони:
1 — голова; 2 — рот; 3 — щупальце; 4 — нога; 5 — мантія; 6 — адаптивні зябра
Нервова система в різних груп черевоногих має різну будову. У нижчих форм гангліїв або немає, або вони слабо виражені, і нервова система складається зі стовбурів, на яких рівномірно розташовані нервові клітини, чим нагадує нервову систему хітонів. Наприклад, у Haliotis (Prosobranchia) є широкий слабо диференційований церебральний тяж, від якого відходять нерви до органів чуття голови, уздовж ноги тягнуться два педальні стовбури, але вже є відокремлені парні парієтальні та один вісцеральний ганглії.
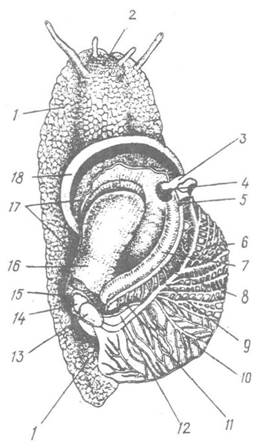
Рис. 67. Helix pomatia з розтятою мантійною порожниною (вигляд зі спинної сторони):
І — нога; 2 — голова; 3, 4, 5 — дихальний, анальний та видільний отвори; 6 — легеневі судини; 7 — пряма кишка; 8 — вторинний сечовід; 9 — легенева вена; 10 — мантія; 11 — первинний сечовід; 12 — нирка; 13 — передсердя; 14 — шлуночок; 15 — перикардій; 16 — дно мантійної порожнини; 17, 18 —лінія розрізу мантії та її край
У більшості черевоногих замість стовбурів утворюються парні ганглії. Такий тип нервової системи називається розкидано-вузловим.
Загальний план будови нервової системи черевоногих наведено на рис. 68. У голові над глоткою розташовані церебральні ганглії, з’єднані між собою церебральною комісурою. У передній частині ноги лежать педальні ганглії, з’єднані між собою під глоткою педальною комісурою, а з церебральними гангліями — конективами. Від церебральних гангліїв відходять довгі стовбури — плевровісцеральні конективи з трьома парами гангліїв: плевральними, парієтальними та вісцеральними; останні можуть з’єднуватися в один непарний. Крім названих, є ще додаткові ганглії, які іннервують окремі органи, наприклад букальний, що іннервує глотку. У всіх Prosobranchia та деяких нижчих представників інших класів плевровісцеральні конективи перехрещуються так, що лівий парієтальний ганглій переміщується на правий бік тіла, а правий — на лівий. Таке явище зветься хіастоневрією (перехрестом). У більшості легеневих та задньозябрових молюсків перехрест конектив зникає, але права плевровісцеральна конектива буває коротша за ліву. У вищих черевоногих (більшість Pulmonata) усі ганглії концентруються навколо глотки, і плевровісцеральні конективи майже не виражені, ніякого перехресту між ними не існує. Так, у виноградного слимака Helix pomatia (Pulmonata) центральна нервова система складається з кільця, яке оточує стравохід (рис. 69); над стравоходом розташовані церебральні ганглії, а під ним —сім тісно зближених гангліїв: пари педальних і плевральних, а також паліальний, парієтальний та абдомінальний. Від них відходять довгі нерви до різних частин тіла.
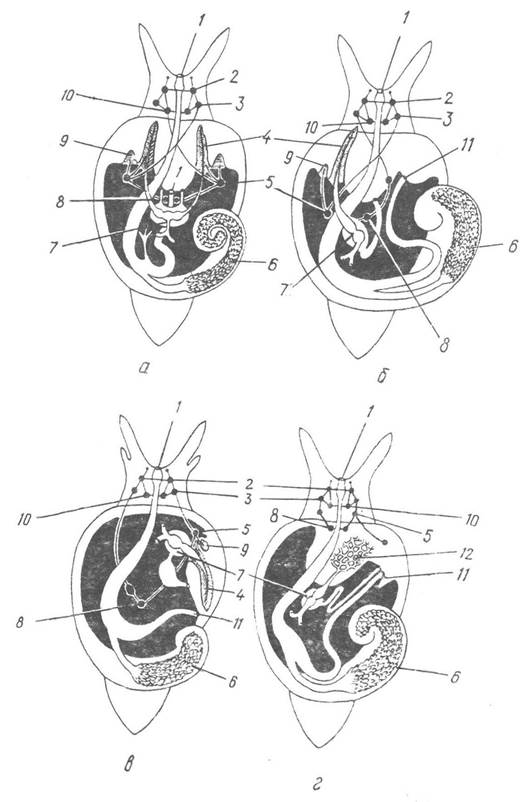
Рис. 68. Схема планів будови черевоногих:
а, б — Prosobranchia з двома та одним ктенідіями відповідно: в — Opisthobranchia; г — Pulmonata: 1 — рот; 2, 3 — церебральний та плевральний ганглії; 4 — ктенідій; 5 — парієтальний ганглій; 6 — печінка; 7— перикардій; 8 — вісцеральний ганглій; 9 — осфрадій; 10 — педальний ганглій; 11 — анус; 12 — легеня
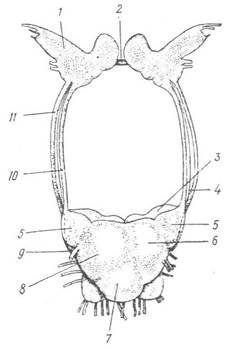
Рис. 69. Центральна нервова система Helix pomatia:
1 — церебральний ганглій; 2 — церебральна комісура; 3 — педальний ганглій; 4 — нерв с тато циста; 5, 6, 7, 8 — плевральний, парієтальний, абдомінальний та паліальний ганглії; 9—статоцист; 10, 11 — цероброплевральна та церобропедальна конективи
Крім центральної нервової системи, у нозі черевоногих залишається периферичне дифузне нервове сплетення, яке міститься в шкірі, а також нервове сплетення у внутрішніх органах (ентодермальне за походженням).
До складу центральної нервової системи Gastropoda входять також нейросекреторні клітини, які виділяють гормони. Вони розташовані в різних гангліях, але найбільше їх у церебральних.
Ендокринні механізми молюсків вивчено значно менше, ніж членистоногих. Відомо, що гормони, які виділяють нейросекреторні клітини, регулюють дозрівання статевих продуктів, водний обмін тощо.
Черевоногі мають різноманітні, добре розвинені органи чуття.
Чуття дотику зосереджено переважно на щупальцях, бічних губах, краях мантії, меншою мірою — на ділянках шкіри, які не прикриті черепашкою.
Органами хімічного чуття є осфрадії та губні щупальця. Осфрадії розташовані біля основи ктенідіїв; якщо є один ктенідій, то відповідно буває один осфрадій.
За будовою він схожий на зябру, тільки менших розмірів, і має форму видовженого валка, по обидва боки якого лежать 90—150 листочків. Поверхня кожного листочка вкрита епітелієм, більша частина якого чутлива. Всередині валка міститься скупчення нервових клітин, від якого відходять нерви до листочків, де утворюють вільні нервові закінчення. Осфрадії іннервуються від парієтальних гангліїв; вони визначають придатність води, що надходить до мантійної порожнини, для дихання. У легеневих молюсків осфрадіїв немає.
Органами смаку та нюху також є передня пара головних щупалець. Спеціальними дослідженнями доведено, що хімічне чуття мають не тільки щупальця, а й шкіра голови та ноги, але найбільшою мірою — це ділянка шкіри, що вистилає вхід до мантійної порожнини.
Усі Gastropoda мають органи рівноваги — статоцисти. Вони лежать на поверхні педальних гангліїв, але іннервують- ся від церебральних. Звичайно це пара замкнених пухирців, епітелій яких складається з війчастих і чутливих клітин, останні іноді утворюють на стінці пухирця особливе скупчення — слухову пляму. Пухирець заповнює рідина, в якій плавають 1—100 конкрецій (слухові камінці). Різне положення їх у статоцистах та натискання то на одну, то на іншу чутливі клітини дає змогу тварині орієнтуватись у просторі.
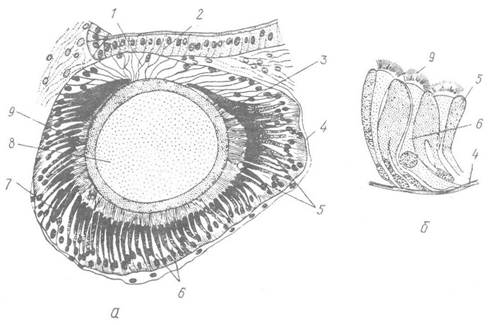
Рис. 70. Око Helix pomatia:
а — зріз через око; б — ділянка сітківки при великому збільшенні; 1, 2 — внутрішня та зовнішня рогівки; 3 — склоподібне тіло; 4 — сполучнотканинний базальний шар; 5, б — пігментні та зорові клітини; 7 — сітківка; 8 — кришталик; 9 — світлочутливі палички-
Органами зору черевоногих є пара очей, розташованих на голові біля основи або на кінчиках задньої пари щупалець. Будова очей різноманітна. У малорухливих видів, наприклад морських блюдечок (Patella), очі найпримітивніші серед черевоногих. Це широко відкритий вгин покривів, дно якого утворює сітківку, або ретину. Остання складається із зорових та опорних клітин. Зовнішня частина ретинального шару утворює темну пігментовану зону, над якою розташований шар світлочутливих паличок, пов’язаних із зоровими (ретинальними) клітинами. Опорні клітини виділяють на поверхні сітківки шар кутикули. Від базальних кінців зорових клітин відходять нервові відростки, що утворюють зоровий нерв, який пов’язаний із церебральним ганглієм. Значно складнішу будову має око виноградного слимака. Воно побудоване за типом очного пухиря (рис. 70). Порожнина його зайнята сферичною лінзою —кришталиком; між ним і стінкою пухиря є шар склоподібного тіла. Око прилягає до епітелію щупальця, який у цьому місці прозорий і зветься зовнішньою рогівкою. Передня стінка очного пухиря пігментована й прозора — це внутрішня рогівка. Задня й бічні стінки ока пігментовані —це сітківка (ретина). Вона складається з двох типів високих клітин, які чергуються одна з одною. Одні з них — пігментні, інші — зорові. Останні на зовнішніх кінцях мають чутливі палички, а базальних —відростки, які утворюють зоровий нерв.
Статева система у черевоногих має різну будову. Prosobranchia здебільшого роздільностатеві, a Puhnonata і Opisthobranchia — гермафродити. Статева залоза майже завжди одна: у роздільностатевих — це яєчник або сім’яник, а в гермафродитів — гермафродитна залоза, в якій утворюються і яйцеклітини, і сперматозоїди. Нижчі черевоногі не мають спеціальних статевих проток, і сіатева залоза відкривається в них у праву нирку. Так, у роздільностатевих морських блюдечок (Patella) непарний сім’яник або яєчник у період розмноження розростається і заповнюється статевими продуктами, які виводяться в порожнину правої нирки через тимчасовий прорив її стінки, а звідти через видільну протоку викидаються назовні; запліднення в них зовнішнє.
Гонади деяких Prosobranchia мають власні вивідні протоки простої будови: у самця — це сім’япровід та копулятивний орган, розташований поблизу від статевого отвору; у самиці — яйцепровід, який може утворювати місцеве розширення — матку, а також бічний виріст — сім’яприймач.
Статеві протоки гермафродитних Pulmonata та Opisthobranchia набагато складніші. Наприклад, у виноградного слимака (див. рис. 60) є одна гермафродитна залоза, яка розташована між частками печінки. Вона продукує і яйцеклітини, і сперматозоїди. Від гермафродитної залози відходить гермафродитна протока, в яку впадає канал особливої білкової залози. Далі вона розширюється і перетворюється на широку складчасту трубку — сім ’яяйцепровід. На поперечному розрізі через нього видно, що його внутрішня порожнина поділена на широку частину, через яку проходять яйцеклітини, і вузенький жолобок, що проводить сперматозоїди. Далі ця спільна протока поділяється на два самостійні канали: яйцепровід і тонший сім’япровід. Яйцепровід сполучається довгим каналом із круглим мішечком — сім’яприймачем. Далі яйцепровід переходить у товстостінну піхву, в яку, крім того, відкриваються два пучки пальцеподібних залоз та товстий великий мішок любовної стріли. Піхва відкривається в статеву клоаку. Сім’япровід переходить у мускулястий чоловічий копулятивний орган, біля основи якого впадає трубчастий придаток — джгут (бич). Копулятивний орган закінчується в статевій клоаці, але при копуляції він випинається назовні.
Гермафродитна залоза виробляє як яйцеклітини, так і сперматозоїди, що надходять спочатку в спільну гермафродитну протоку. Білкова залоза виділяє білок, який обволікає запліднені яйцеклітини і є поживною речовиною для зародка. Джгут виділяє клейку речовину, яка склеює сперматозоїда в сперматофор. Сім’яприймач слугує для зберігання чужої сперми, одержаної при паруванні. Любовна стріла під час копуляції випинається через статеву клоаку і слугує для подразнення партнера, втикаючись у його шкіру.
У виноградного слимака під час копуляції кожна особина виконує роль і самця, і самиці, при цьому вони обмінюються сперматофорами. Після цього сперматофори із сперматозоїдами проникають у сім’яприймачі. Запліднені яйця оточуються секретом білкової залози та захисною оболонкою. Слимак викопує в грунті ямку і відкладає туди яйця.
У деяких видів черевоногих у гермафродитній залозі розвиваються по черзі то яйцеклітини, то сперматозоїда, і тому такі слимаки в якийсь певний час бувають або самцями, або самицями.
Більшість черевоногих відкладають яйця в коконі. Водяні форми іноді оточують яйце драглистою масою у вигляді стрічок, шнурів тощо, а наземні закопують яйця в землю. Бувають випадки піклування про нащадків: яйця виношуються молюсками на поверхні тіла або в мантійній порожнині. Нарешті, трапляються і живородні форми, такі, наприклад, як прісноводні живородки Viviparus, в яких запліднені яйця розвиваються в яйцепроводі, а назовні виходить уже сформована молодь.
Яйця черевоногих найчастіше мають мало жовтка. Дробіння в них проходить за спіральним типом і багато в чому нагадує дробіння поліхет. Гаструляція відбувається шляхом інвагінації або епіболії, або вростанням всередину бластули щільного зачатка. Бластопор набуває щілиноподібної форми і замикається ззаду наперед; на місці його переднього краю залишається отвір, який перетворюється на ротовий.
У примітивніших Gastropoda (наприклад, у Patella) гаструла перетворюється на личинку трохофору (рис. 71, а), яка має тім’яний орган з китицею війок, прототрох і наскрізний кишечник. Стадія трохофори нетривала і властива не всім черевоногим. Вона невдовзі перетворюється на велігера (рис. 71, бв), характерними ознаками якого є наявність паруса, зачатків черепашкової залози, ноги та радули. Парус має вигляд 2- або 3-лопатевої шкірної складки, по краю якої проходять прототрох і метатрох з харчовою борозенкою поміж ними. Черепашкова залоза закладається як впинання ектодерми на спинній стороні личинки, на дні якого виділлється органічна речовина черепашки (конхіолін), а потім вона вивертається, утворюючи ніжну личинкову черепашку. Згодом на органічній основі черепашки відкладаються вапнякові шари. Краї черепашкової залози перетворюються на край мантії. У вигляді вип’ячування шкірних покривів на черевній стороні тіла з’являється нога.
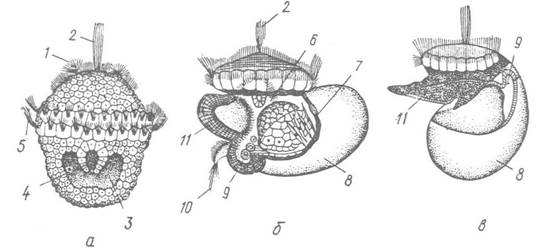
Рис. 71. Розвиток Patella (Prosobranchia):
а — трохофора; б — велігер перед закручуванням; в — після нього; 1 — війки верхньої півкулі; 2 — тім’яний пучок війок; 3 — мезодермальні смужки, що просвічують крізь покриви; 4 — рот; 5 — прототрох; 6 — кишечник; 7 — нутрощевий мішок; 8 — черепашка; 9 — мантія; 10 — війки задньої частини тіла; 11 — зачаток ноги
Після більш-менш тривалого періоду планктонного життя велігер опускається на дно і завершує метаморфоз; личинкові органи (шкірні покриви, парус тощо) дегенерують. У ході метаморфозу білатеральносиметрична личинка набуває асиметрії, характерної для дорослих форм.
У більшості черевоногих стадія трохофори формується під захистом яйцевих оболонок, з яких виходить вже велігер. В усіх наземних та прісноводних слимаків, а також частини морських з яйця виходить цілком сформований молюск, який значно менший за дорослого і має черепашку всього з одним-двома обертами. Ріст тіла слимаків і збільшення розмірів черепашки відбувається шляхом поступового утворення нових обертів або розростанням в ширину одного чи двох останніх обертів. Саме такий спосіб характерний для видів із широкою ковпачкоподібною черепашкою.
План будови дорослих черевоногих відрізняється від плану будови личинки. У Gastropoda, на відміну від хітонів, соленогастрів та двостулкових, відбувається посилений ріст спини при малому рості в довжину. Це призводить до відносного зближення рота й ануса та утворення петлі кишечника («анопедіальний вигин»). На спині утворюється горб — нутрощевий мішок, в який втягується кишечник (рис. 71, б). Цей горб вкривається черепашкою і для більшої компактності закручується спіраллю.
Друга перебудова полягає в торсійному процесі, який відбувається в усіх черевоногих (рис. 72). Це —поворот нутрощевого мішка разом із черепашкою, яка його вкриває, та мантійним комплексом органів відносно голови й ноги проти годинникової стрілки (якщо дивитись зі спини) на 180°. У результаті мантійний комплекс органів опиняється біля переднього кінця тіла; зябра верхівками спрямовані вперед, а закруток черепашки обернений назад. Наслідком торсійного процесу є перехрест плевровісцеральних конектив (хіастоневрія).
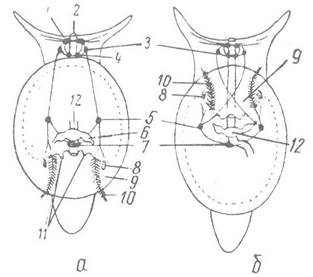
Рис. 72. Схема розташування мантійною комплексу органів черевоногого молюска:
а, б — перед та після його закручування; 1 — церебральний ганглій; 2 — рот; 3, 4, 5 — плевральний, педальний та парієтальний ганглій; 6 — передсердя; 7 — вісцеральний ганглій; 8 — осфрадій; 9 — мантійна порожнина; 10 — ктенідій; 11 — видільні отвори; 12 — шлуночок серця
Торсійний процес у різних підкласах проходить по-різному. У найпримітивніших Prosobranchia (наприклад, Patella) він відбувається на стадії велігера протягом кількох хвилин, коли личинки крутять свій нутрощевий мішок у різних напрямах, а потім повертають його останній раз, і таке положення фіксується назавжди. У більшості черевоногих торсійний процес відбувається під час ембріонального розвитку шляхом нерівномірного росту окремих частин зародка, а з яйця виходить вже стадія з хіастоневрією. У результаті торсійного процесу виникає асиметрія лише в стебельці, тобто тій частині тулуба, яка з’єднує голову й ногу з нутрощевим мішком.
Інші прояви асиметрії пов’язані з асиметрією самого нутрощевого мішка, який повторює оберти турбоспіралі черепашки, та асиметрією вустя черепашки, яка виникає внаслідок зміни способу її носіння (рис. 73). Більшість слимаків із турбоспіральною черепашкою носить її в косому положенні: верхівка черепашки піднята вгору и відведена дещо назад, але розташована не на середній лінії тіла, яка проходить через білатеральносиметричні голову й ногу, а праворуч від неї. Через це вустя черепашки лежить навскіс, і рух води, що надходить до мантійної порожнини для дихання, є нерівномірним: до ширшої лівої частини вода надходить інтенсивніше, ніж до правої. Внаслідок цього у більшості Prosobranchia з турбоспіральною черепашкою правий ктенідій або менший за лівий, або зовсім редукований. Відповідно редукується й праве передсердя, до якого йде кров з цього ктенідія. Описані перетворення відбуваються в індивідуальному розвитку приховано, шляхом нерівномірного росту органів правої і лівої сторін.
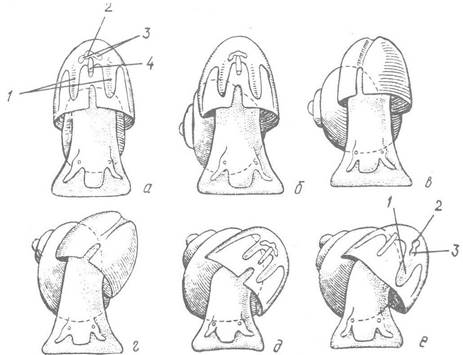
Рис. 73. Схема аміни способу носіння черепашки при переході її від плоско спіральної до турбоспіральної форми (вигляд зверху):
а—в — висування верхівки черепашки вбік і утворення турбоспіралі; г—е — зміна способу носіння черепашки та утворення асиметрії мантійного комплексу; 1 — ктенідії; 2 — шлуночок серця; 3 — передсердя; 4 — задня кишка
В Opisthobranchia мантійна порожнина напрямлена не вперед, а назад, і лежить на правій стороні тіла, хіастоневрії немає. Вважають, що це є наслідком повороту черепашки в протилежному, ніж при торсійному процесі, напрямку, тобто за годинниковою стрілкою, у результаті чого плевровісце- ральні конективи розкручуються. У Pulmonata, хоча мантійний комплекс органів і напрямлений уперед, але хіастоневрії теж немає, проте це пов’язано з укороченням або зникненням конектив і концентрацією всіх гангліїв навколо глотки (див. рис. 68).
Черевоногі є складовими багатьох біоценозів морів, океанів, прісних водойм та суходолу. Багато видів черевоногих поїдається рибами, амфібіями, рептиліями, птахами та ссавцями. На суходолі ними живляться також комахи. У порожні черепашки водяних слимаків заселяються поліхети, губки, кишковопорожнинні, личинки комах, а черепашки наземних слимаків використовують мух та ос для відкладання яєць.
Багато видів черевоногих використовується людиною в їжу (виноградний слимак, галіотіси, букцинуми, літорини тощо), а також для виготовлення прикрас, ґудзиків, амулетів, для інкрустаційних робіт, особливо ціняться черепашки з розвиненим перламутровим шаром.
Негативне значення черевоногих полягає в тому, що деякі з них є проміжними хазяями трематод — паразитів людини та свійських тварин (ставковики, бітінії).
Існують різні варіанти системи класу Gastropoda. Традиційно їх поділяють на три підкласи: Передньозяброві (Ргоsobranchia), Задньозяброві (Opisthobranchia) та Легеневі (Pulmonata), хоча останнім часом Передньозяброві розбивають на кілька самостійних підкласів. Проте нова система ще не є загальновизнаною, тому наводимо традиційну систему.