ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 3 - Г.Й. Щербак - 1997
ТИП МОЛЮСКИ, АБО М’ЯКУНИ (MOLLUSCA)
КЛАС ГОЛОВОНОГІ (CEPHALOPODA)
Головоногі — виключно морські організми, найбільше поширені в тропічних та субтропічних водах, але мешкають і в помірних, і в полярних морях. Вони трапляються скрізь від поверхні до найбільших глибин. Серед них є бентосні (придонні), нектонні (ті, що активно плавають у товщі води) та планктонні форми. Головоногі можуть жити лише в океанах і морях із солоністю не нижчою, ніж 33 %о, тому їх немає ні в Чорному, ні в Азовському морях. Тільки деякі прибережні види кальмарів - лолігінід, що мешкають біля берегів Центральної Америки, можуть витримувати тимчасове опріснення. Усього описано близько 650 видів головоногих, проте загальна їх кількість може бути більшою, оскільки ще не завершено дослідження глибоководної фауни, де в останні десятиріччя знаходять нові, ще не відомі науці види.
Головоногі — найбільш високоорганізована група молюсків і одні з найбільш високорозвинених безхребетних. Це досить давня група молюсків, відома з раннього палеозою. Фауна викопних головоногих значно багатша від сучасної — вона налічує близько 10 тис. видів.
За розмірами головоногі бувають досить різними, серед них є дуже дрібні форми, довжина мантії яких менша за 1 см (деякі каракатиці), а є й велетні, такі, як велетенський кальмар Architeuthisdux, довжина якого разом зі щупальцями може сягати 18 м. Це виключно хижі, надзвичайно рухливі тварини.
Головоногі — білатеральносиметричні тварини із зовнішньою (підклас Nautiloidea) або внутрішньою (підклас Соleoidea) рудиментарною черепашкою. Тіло їх складається з голови та тулуба, покритого мантією. У каракатиць тулуб сплющений, у кальмарів — циліндричний, загострений на задньому кінці, у восьминогів —мішкоподібний (рис. 90).
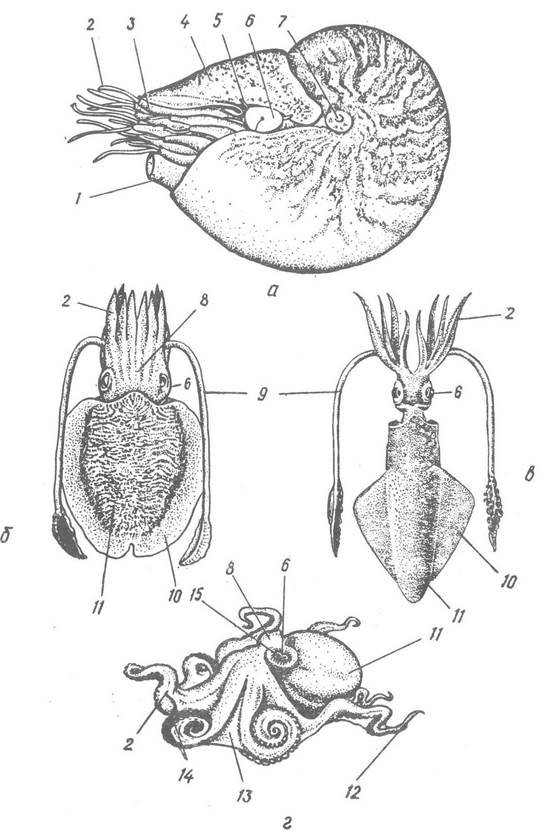
Рис. 91. Зовнішня будова представників класу Cephalopoda
Велика голова, як правило, добре відокремлена від тулуба і має очі, вінець кінцівок (рук, або ніг), що оточують ротовий отвір, та лійку. У колеоідей 8 або 10 кінцівок: у кальмарів та каракатиць завжди 8 рук та пара щупалець; у восьминогів тільки 8 рук. Наутилоідеї мають близько 90 рук.
Внутрішня поверхня рук головоногих (за винятком Nautilus) вкрита присосками, які розташовані одним—чотирма, рідко більше, поздовжніми рядами. Найкрупніші присоски розташовані посередині рук або ближче до їх основи, а найменші — на кінцях рук.
Кальмари та каракатиці, крім восьми рук, мають ще пару ловецьких щупалець, які складаються із стебла без присосків та розширеної булави з присосками (рис. 90).
У кальмарів та каракатиць присоски містяться на коротких ніжках і озброєні роговими кільцями з гладенькими або зазубреними краями; у деяких океанічних кальмарів вони перетворюються на міцні гачки, що нагадують кошачі кігті, які допомагають їм хапати й утримувати здобич (рис. 91). У вампіроморф та восьминогів присоски мають простішу будову: вони позбавлені ніжок, рогових кілець та гачків. У вампіроморф та плавцевих восьминогів між присосками розташовані коротенькі вусики, які тягнуться правильними рядами обабіч кожного ряду присосків і виконують функцію органів дотику.
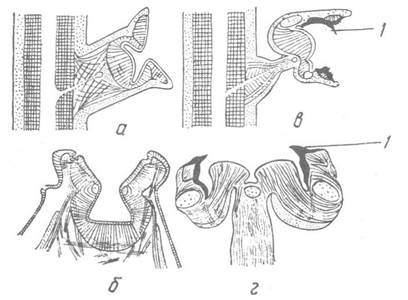
Рис. 91. Схема будови присосків:
а, б — восьминогів; в — кальмарів; г — каракатиць; 1 — зубці
На нижній стороні голови міститься лійка — конічна, звужена спереду трубка. Через неї викидається вода з мантійної порожнини при диханні та реактивному плаванні, виводяться назовні екскременти, сеча, чорнильна рідина, яйця. Бічні сторони лійки зростаються з мантією або з’єднуються з нею за допомогою замикальних хрящів.
Тулуб з усіх боків вкритий мантією. На спині мантія утворює покриви самого тулуба, а на черевній стороні між нею та стінкою тіла залишається мантійна порожнина, яка сполучається із зовнішнім середовищем щілиноподібним черевним отвором.
Для замикання черевної щілини в каракатиць та кальмарів є особливе пристосування у вигляді півмісяцевих ямок на основній частині лійки, відповідно до яких на внутрішній поверхні мантії лежать два великих, укріплених хрящем, горбки (рис. 92). Ці утвори діють за принципом застібки-кнопки.

Рис. 92. Sepia officinalis з розтятою мантійною порожниною:
1 — дистальне розширення ловецького щупальця; 2 — його присоски; 3 — ловецьке щупальце; 4 — руки; 5 — рот; 6 — передній отвір лійки; 7 — передній та 8 — задній відділи лійки; 9 — анальний отвір; 10 — ктенідій; 11 — видільний отвір; 12 — статевий отвір; 13 — зяброва вісь; 14 — отвір нідаментальної залози; 15 — протока чорнильного мішка; 16 — чорнильний мішок; 17 — плавець; 18 — товща мантії; 19 — лопаті нідаментальної залози; 20 — мантія; 21 — м’яз, що відтягує лійку; 22 — горбок замикального апарату; 23 — мантійний ганглій, що просвічує крізь покриви; 24 —ямка замикального апарату; 25 — нюшна ямка
У мантійній порожнині міститься мантійний комплекс органів: анальний отвір, по боках якого розташовані видільні й статеві (один або два) отвори та пара (у наутилуса — дві пари) пірчастих ктенідіїв. Крім того, у самиць поблизу від статевого отвору до мантійної порожнини відкриваються протоки двох парних та однієї непарної нідаментальних залоз (див. далі).
Мантія нектонних (активно плаваючих) кальмарів та каракатиць має таку будову: зовні вона вкрита шкірою, під нею лежить зовнішня підшкірна оболонка — туніка з численними колагеновими волокнами і тонким шаром поздовжніх м’язів, потім власне мантія — м’язовий шар, глибше — тонка внутрішня туніка (також із волокнами та нечисленними поздовжніми м’язами) і тонка шкірна вистилка мантійної порожнини. Власне мантія має товщину 2—3 см у активних нектонних видів та близько 0,5—1 см у менш рухливих. Вона складається з шарів кільцевих та радіальних м’язів, які чергуються. До складу м’язових шарів входять також колагенові волокна. Поперемінне скорочення радіальних і кільцевих м’язів забезпечує розширення та скорочення мантії, а еластичність колагенових волокон — відновлення форми мантії після м’язового скорочення.
У напівпланктонних та планктонних головоногих м’язи більш-менш редуковані, мантія напівпрозора і за консистенцією нагадує дзвін медузи. Товста стінка мантії планктонних океанічних кальмарів заповнена вакуолями з розчином NH4Cl.
На задньому кінці або посередині мантії в більшості колеоідей міститься пара плавців — основний орган при повільному плаванні. Вони завжди мускулясті, навіть у медузоподібних головоногих. Плавці можуть мати вигляд вузької кайми, яка тягнеться по боках мантії (у каракатиць-сепіїд та деяких кальмарів, наприклад Sepioteuthis), або бути ромбічними, серцеподібними, округлими тощо. У звичайних восьминогів плавців немає.
Черепашка у більшості головоногих рудиментарна або її зовсім немає і добре розвинена лише в кораблика (Nautilus, рис. 93). Вапнякова зовнішня черепашка наутилуса закручена в одній площині на спинну сторону і повернена закрутком уперед. Її порожнина поділена поперечними перетинками на ряд камер. Тіло тварини міститься лише в останній, найбільшій, камері, а решта камер заповнені газом і частково рідиною. Посередині кожної перетинки є по невеличкому отвору. Через ці отвори всі камери пронизує тонкий циліндричний відросток — сифон, який є виростом нутрощевого мішка.
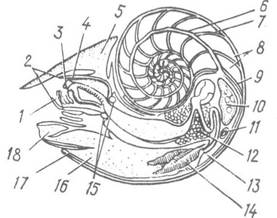
Рис. 93. Розріз через тіло самиці Nautilus pompilius:
1 — нижня шелепа; 2 — руки; 3 — радула; 4 — верхня щелепа; 5 — каптур; б — сифон; 7 — септа; 8 — камери; 9 — задній край мантії; 10 — яєчник; 11 — серце; 12 — травна залоза; 13 — анус; 14 — ктенідії; 15 — ганглії; 16 — мантійна порожнина; 17—мантія; 18 — лійка
У решти головоногих залишається лише рудимент черепашки, яка обгортається мантією і стає внутрішньою. У каракатиць черепашка має вигляд великої видовженоовальної вапнякової пластинки (рис. 94, а, б). Зрозуміти її будову можна, лише знаючи будову черепашки викопних Cephalopoda — белемнітів. В останніх черепашка була багатокамерною, як у наутилуса. Вона складалася з прямого конуса — фрагмокона, поділеного поперечними перетинками (септами) на ряд камер (рис. 94, в). Септи пронизувались поздовжньою сифональною трубкою, в якій містився сифон — м’язистий відросток нутрощевого мішка. Спинна стінка фрагмокона була витягнена вперед у вигляді тонкого листоподібного рогового виросту —проостракума. Крім того, фрагмокон був вкритий масивним вапняковим футляром у вигляді загостреного ззаду конуса — рострума. Здебільшого від черепашок белемнітів залишаються саме ці кінцеві футляри, які у народі називають «чортовими пальцями».
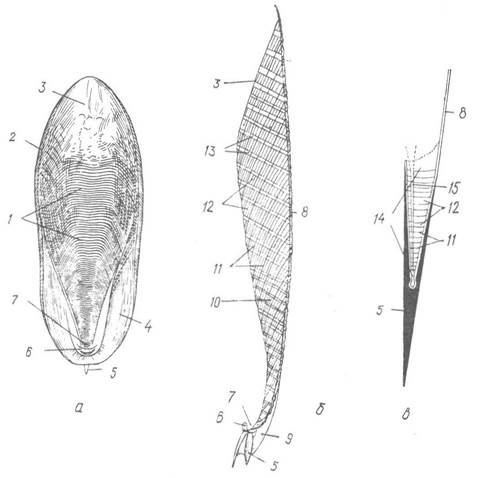
Рис. 94. Будова черепашок представників підкласу Coleoidea:
а — черепашка Sepia officinalis з черевної сторони; б — її медіальний розпил; в — схема поздовжнього розпилу черепашки белемніта;
1 — зближені перетинки спинної сторони фрагмокона; 2 — бічний край проостракума; 3 — поверхня наймолодшої септи; 4 — задній край проостракума; 5 — рострум; 6 — рудимент черевної стінки; 7 — сифональна лійка; 8 — проостракум; 9 — потовщення зовнішньої конхіолінової пластинки; 10 — додаткові конхіолінові пластинки; 11 — септи; 12 — камери; 13 — опорні вапнякові стовпчики; 14 — фрагмокон; 15 — сифональна трубка
Черепашка каракатиці, яка називається сепіоном, має всі ті самі основні елементи, що й черепашка белемнітів, але дуже видозмінена порівняно з нею. Проостракум представлений опуклою і широкою спинною пластинкою. До нього з черевної сторони прилягають сильно зближені та скошені септи спинної частини фрагмокона, а черевна його частина майже повністю зникає. Проміжки між сусідніми септами відповідають камерам черепашки белемнітів; їх перетинають численні вапнякові стовпчики, перпендикулярні до площини перетинок. Отже, спинна частина фрагмокона каракатиці має вигляд товстої, пористої пластинки, а черевна його частина майже зовсім зникла, лише на задньому кінці зберігається редукована черевна стінка у вигляді невеличкої лієчки, порожнина якої є задньою ділянкою сифона, що збереглася. Сифональна трубка, порівняно з такою белемнітів, коротенька, лійкоподібно розширена. Дуже редукований рострум має форму невеличкого шипа на задньому кінці фрагмокона. Внутрішня черепашка каракатиць захищає внутрішні органи і править за гідростатичний апарат; вона, крім того, є опорою для м’язів.
Серед сучасних колеоідей лише глибоководний вид Spirilla spirula має спіралько закручену внутрішню черепашку, що складається з 25—35 камер, крізь які проходить сифон (рис. 95); вона є для спірули гідростатичним апаратом, який дає змогу їй жити на глибині 1750 м і підніматись у поверхневі шари води до 100—300 м.
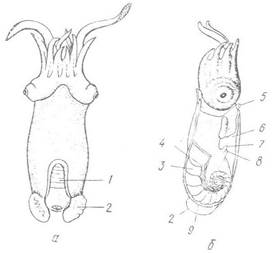
Рис. 95. Spirula spirula (ряд Sepiida) із спіральною багатокамерною черепашкою:
а — вигляд зі спинної сторони; б — сагітальний розріз; 1 — ділянка черепашки, вкрита мантією; 2 — плавець; 3 — черепашка; 4 — сифон; 5 — лійка; 6 — мантійна порожнина; 7 — анус; 8 — видільний отвір; 9 — орган свічення
У вампіроморф та кальмарів від черепашки залишається лише спинний роговий листок — гладіус («гладіаторський щит»). Це —вузька пероподібна, ланцетоподібна або голкоподібна рогова (але з домішкою хітину) пластинка, що лежить на спині під шкірою (рис 96). У донних восьминогів-октоподид гладіус редукований до двох тонких хрящових паличок, а у вищих сепіолід та пелагічних восьминогів його зовсім немає. У більшості плавцевих восьминогів від черепашки залишається хрящова пластинка сідло- або підковоподібної форми, яка підтримує плавці.

Рис. 96. Гладіус кальмарів:
а — Todarodes pacificus; б — Loligo sp.; 1 — стовбур; 2 — перо; 3 — кінцевий конус
Рудиментарна черепашка всіх колеоідей вільно лежить всередині замкненого епітеліального мішка, епітелій стінок якого виділяє гладіус.
Виключно своєрідну черепашку мають самиці пелагічного восьминога-аргонавта, або паперового кораблика (Argonauta argo). Це зовнішня, дуже тонкостінна, ніби пергаментна, спірально закручена на спину черепашка, яка не поділена на камери і ніде щільно не прилягає до тіла (рис. 97). Вона утворюється лише в самиць на 10—12-й день після народження. Її виділяє не спинна черепашкова залоза, як справжню черепашку, а епітелій однієї пари розширених спинних рук. Це вторинний утвір, що не має нічого спільного із справжніми черепашками і слугує для виношування яєць.
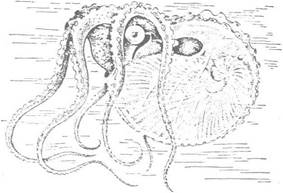
Рис. 97. Аргонавт (Argonauta argo, ряд Octopoda)
На відміну від інших молюсків, у головоногих, крім черепашки, є ще добре розвинений внутрішній скелет, який складається з хряща, подібного за будовою до хряща хребетних. Найбільше він розвинений у Coleoidea. Це головна хрящова капсула, яка оточує центральну нервову систему; від неї відходять вирости, що оточують очі та статоцисти (рис. 98). Це утворення подібне до черепа хребетних. Є також опорні хрящі в основі щупалець, усередині плавців та в замикальному апараті мантії — «кнопках». Хрящові утворення головоногих мають мезодермальне походження.
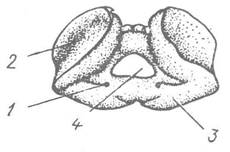
Рис. 98. Головна капсула Octopus dofleini:
1 — отвір для кровоносної судини; 2 — заглиблення для ока; 3 — капсула для с тато циста; 4 — задній отвір капсули
Шкіра головоногих складається з одношарового циліндричного епітелію, під яким залягає сполучна тканина. Шкірний епітелій містить багато слизових клітин. Слиз робить тіло головоногих слизьким, що полегшує їм рух у воді. Сполучнотканинний шар містить м’язові волокна та пігментні клітини; під ним лежить зовнішня туніка.
Пігментні клітини бувають двох типів: хроматофори та іридоцити. Хроматофори — клітини, що містять пігмент. Це великі зірчасті клітини, до яких прикріплюються радіально розташовані навколо них м’язові клітини; до останніх підходять нервові закінчення (рис. 99). При скороченні м’язів хроматофори розтягуються, збільшуючись у десятки разів, а при їх розслабленні вони набувають початкових розмірів. Хроматофори містять чорні, коричневі, червоно-бурі, оранжеві та жовті пігменти. Іридоцити — це сплющені овальні клітини, в яких містяться блискучі тільця — іридосоми, які відбивають та заломлюють світло, розкладаючи його на різні кольори спектра. Різні співвідношення пігментного забарвлення з ефектом, який створюють іридоцити, дає незліченну різноманітність відтінків, у які може забарвлюватися головоногий молюск.

Рис. 99. Будова сполучнотканинного шару шкіри каракатиці :
1 — м’язові пучки; 2 — іридоцит; 3 — м’язові клітини хроматофора; 4 — хроматофор; 5 — шкірна кровоносна судина; 6 — ядро м’язової клітини
Зміна забарвлення регулюється головним мозком, а також сіткою нервових клітин, розташованих навколо хроматофорів. Зміна забарвлення пов’язана із сигналами, які надходять від органів зору. Якщо осліпити восьминога на одне око, він втрачає здатність змінювати забарвлення відповідної сторони тіла. Каракатиці та восьминоги за допомогою хроматофорів маскуються під колір та малюнок субстрату. Це маскування використовується як для захисту, так і для підстерігання здобичі: тварина, заховавшись між придонними предметами, робиться непомітною. Крім того, різкою зміною забарвлення молюск відлякує ворогів.
У шкірі головоногих є також особливі органи, що світяться — фотофори. Будова фотофорів дуже різноманітна. У найгфостішому випадку це скупчення фотоцитів, пронизане кровоносними судинами та оточене оболонкою з вакуолізо- ваних клітин. Такі фотофори можуть бути розкиданими по всій шкірі або зібраними в щільні групи. Бувають і значно складніші фотофори. Часто фотофор нагадує автомобільну фару напівсферичної форми. З усіх боків, крім поверхні, що світиться, він вкритий непроникним для світла пігментним шаром, а дно його, як дзеркальний рефлектор, вистелене блискучим шаром. Усередині лежить джерело світла — маса фотоцитів. Зверху «фару» прикриває прозора лінза, а поверх неї — діафрагма, яка складається з шару чорних хроматофорів. Насуваючи на лінзу діафрагму, тварина може регулювати інтенсивність свічення фотофора і навіть повністю його загасити.
Фотофори є в переважної більшості кальмарів, окремих видів каракатиць (Sepia australis) та восьминогів (Octopus ornatus, Tremoctopus violaceus), а також у спірули і вампіротейтіса. У головоногих розрізняють два типи свічення: симбіотичне (бактеріальне) та власне (внутрішньоклітинне). При симбіотичному свіченні світяться бактерії роду Photobacterium, які живуть у залозах, розташованих на чорнильному мішку. Ці органи є в деяких донних каракатиць та кальмарів. Бактерії можуть світитися всередині залози; крім того, слиз із бактеріями, що світяться, може викидатися через лійку назовні.
Власне свічення зумовлене реакцією окислення люциферина атомарним киснем під дією фермента люциферази, яка проходить у цитоплазмі особливих клітин — фотоцитів. Таке свічення властиве переважно пелагічним головоногим.
Фотофори виконують різні функції. З їхньою допомогою тварини відлякують ворогів, а також упізнають одне одного. Крім того, фотофори, наприклад «ліхтарики» на кінцях довгих щупалець кальмара Chiroteuthis (рис. 100), можуть приваблювати здобич. В океанічних кальмарів головною функцією фотофорів, які містяться на черевній стороні тіла, вважають створення вентрального протисвічення. Кальмар, який не світиться, чітко вирізняється на фоні світлого неба, якщо дивитися на нього знизу вдень; якщо він буде тьмяно світитися, то може стані непомітним.
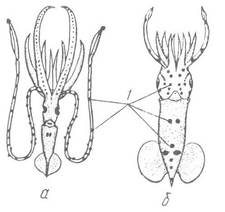
Рис. 100. Схема розташування фотофорів на тілі кальмарів:
а — Chiroteuthis calyx; б — Pterygioteuthis gemmata; в — фотофори
Мускулатура головоногих відрізняється особливо сильним розвитком (рис. 101). Уся мантія містить велику кількість м’язів, які залягають у вигляді суцільного шару. Найбільше розвинені поперечні м’язи. При скороченні вони притискують мантію до черевної стінки тулуба, сприяючи виштовхуванню води через лійку. Дуже добре розвинені кільцеві й поздовжні м’язи рук, а також м’язи присосків. Крім описаних м’язів, які нагадують мускулатуру шкірно-м’язового мішка, є й спеціалізовані, у вигляді могутніх пучків. Це м’язи, що скорочують голову, або рухають лійку; вони віялоподібно відходять від черепашки до лійки й голови.
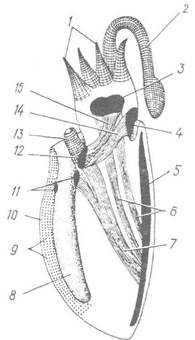
Рис. 101. Схема мускулатури каракатиці (сагітальний зріз):
1, 2 — м’язи рук та ловецького щупальця; 3 — головна капсула; 4 — потиличний хрящ; 5 — черепашка; 6 — ретрактори голови; 7 — м’язи, що втягують лійку; 8 — мантійна порожнина; 9 — перерізані поперечні м’язи мантії; 10 — товща мантії; 11 — хрящі застібки-кнопки; 12 — хрящ лійки; 13 — лійка; 14 — шийний м’яз; 15 — м’яз, що підтримує лійку
Головоногі дуже рухливі тварини. Вчені підрахували, що великі за розміром кальмари можуть розвивати швидкість до 40—55 км/год.
Целом у головоногих, на відміну від інших молюсків, великий: у ньому містяться внутрішні органи. Як і в інших молюсків, він складається зі статевого та перикардіального відділів, але обидва відділи сполучаються між собою протокою, яка називається водоносним каналом. Найбільший целом, навіть без перетяжин між двома відділами, має пекельний кальмар-вампір (ряд Vampyromorpha). У Nautilus статевий целом утворює справжню порожнину тіла, в якій лежать гонада, шлунок та частина кишечника. Він переходить у сифон, який тягнеться всередині закрутки черепашки. Перикардіальний целом, який містить серце, також
великий; обидва відділи сполучаються протокою. У каракатиць та кальмарів у об’ємному статевому відділі целома (рис. 102) розташовані гонада та шлунок; у меншому, перикардіальному, відділі лежать серце, зяброві серця та перикардіальні залози. У восьминогів добре розвинена паренхіма, і целом дуже редукований; серце лежить не в перикардію, а в паренхімі. Розвинена лише статева частина целома, яка представлена порожниною гонади.
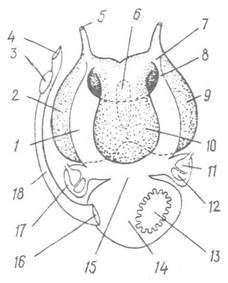
Рис. 102. Целом, видільна та статева системи самиці Sepia officinalis (вигляд зі спинної сторони):
1 — перикардіальний відділ целома; 2 — лівий нирковий мішок; 3 — залоза яйцепроводу; 4 — статевий отвір; 5 — зовнішній отвір лівої нирки; б — проміжок між черевними нирковими мішками; 7—реноперикардіальний отвір; 8 — реноперикардіальний канал; 9 — правий нирковий мішок; 10 — спинний нирковий мішок; 11 — зяброве серце; 12 — перикардіальна залоза; 13 — яєчник; 14 — статевий відділ целома; 15 — сполучення перикардіального та статевого відділів целома; 16 — отвір, що веде із статевого відділу целома до яйцепроводу; 17 — целомічний мішок зябрового серця; 18 — яйцепровід
Основні функції целома головоногих — захист серця, участь у процесах виділення і сольового обміну та у виведенні статевих продуктів. Особливу функцію виконує целом кранхіїд, заповнений розчином хлориду амонію (див. с. 138).
Травна система головоногих досягає високого рівня диференціації (рис. 103, 104). Рот міститься на голові в центрі вінця рук. Він веде в мускулясту глотку, яка має пару міцних рогових щелеп (верхню та нижню), схожих на дзьоб папуги (рис. 105). У глотці є язик з вузькою радулою. За допомогою щелеп молюски вбивають здобич та відривають від неї шматки; цілу здобич вони не ковтають. Зубці радули підхоплюють відкушені шматки їжі та переносять їх до глотки; у восьминогів радула використовується також для просвердлювання черепашок двостулкових молюсків, які є складовою їхнього раціону.

Рис. 103. Загальна схема розташування внутрішніх органів самиці каракатиці:
I — хрящова капсула; 2 — глотка із дзьобом; 3 — мозок; 4 — лійка; 5 — анус; б — протока чорнильного мішка; 7 — пряма кишка; 8 — ктенідій; 9 — мантія; 10 — шлуночок серця;
II — задня аорта; 12 — чорнильний мішок; 13 — статевий відділ целома; 14 — яєчник; 15 — сліпий мішок шлунка (цекум); 16 — перикардіальний відділ целома; 17 — шлунок; 18 — черепашка; 19 — травна залоза (печінка)

Рис. 104. Травна система каракатиці Sepia officinalis:
1 — глотка; 2 — розріз через нервове кільце; 3 — порожнина капсули статоциста; 4 — розрізана головна капсула; 5 — печінкова протока; 6 — анус; 7 — протока чорнильного мішка; 8 — пряма кишка; 9 — тонка кишка; 10 — сліпий мішок шлунка (цекум); 11 — шлунок; 12 — підшлункова залоза; 13 — головна аорта; 14 — печінка; 15 — стравохід; 16 — задня слинна залоза; 17 — слинна протока; 18 — загальна слинна протока

Рис. 105. Дзьоб кальмара:
1,2 — нижня та верхня щелепи
У глотку впадають протоки двох пар слинних залоз. Секрет передніх залоз містить травні ферменти; задні, крім ферментів, часто виділяють отруту, яка паралізує або вбиває здобич (ракоподібних тощо). Від глотки відходить довгий стравохід — встелена кутикулою трубка, яка проходить крізь мозок, тому головоногі змушені розкушувати їжу на дрібні шматки, а не ковтати її цілком. У наутилуса та безплавцевих восьминогів стравохід утворює бічний випин — воло, у наутилуса воло дуже велике, навіть більше за шлунок.
Стравохід відкривається в шлунок, який поділений на дві частини: власне шлунок та сліпий мішок шлунка — цекум. Власне шлунок встелений кутикулою та має мускулясті стінки. Внутрішня поверхня це кума вкрита війчастим епітелієм із великою кількістю слизових клітин. У цекум відкриваються протоки травної залози (печінки).
Печінка велика, часто складається з двох часток. Печінкові протоки оточені численними залозистими придатками, які називаються підшлунковою залозою. У кальмарів та каракатиць це подвійний орган, який встелений двома шарами епітелію. До травної системи належить лише внутрішній шар епітелію, а зовнішній становить частину видільної системи. Тільки у восьминогів підшлункова залоза цілком належить до травної системи. Від шлунка відходить тонка кишка, яка відкривається анальним отвором у передній частині мантійної порожнини.
До ектодермального переднього відділу травної системи належать глотка із слинними залозами та стравохід; до ентодермального середнього — шлунок із цекумом, печінка з підшлунковою залозою та кишка з чорнильним мішком. Ембріологічні дослідження показали, що ектодермальної задньої кишки в головоногих немає.
Головоногі — виключно хижі тварини, які живляться живою здобиччю, лише Nautilus споживає й падаль. Секрет слинних залоз містить ферменти, які розщеплюють вуглеводи та білки. їжа, змочена слиною, потрапляє в шлунок, куди надходять також секрети печінки, підшлункової залози та сліпого мішка з травними ферментами. Мускулатура шлунка забезпечує постійне перемішування їжі й травних соків. У шлунку відбувається початкове травлення, далі напівперетравлена їжа у вигляді емульсії надходить у цекум, а з нього по печінкових протоках — у підшлункову залозу та печінку. У цекумі завершується травлення і починається всмоктування жирів, амінокислот та вуглеводів. Але основним органом всмоктування є печінка, де всмоктується 65— 95 % продуктів травлення. Підшлунковою залозою також всмоктуються вуглеводи та амінокислоти. Крім того, в ній відбувається секреція та осморегуляція.
Печінка виконує дуже різноманітні функції. Вона виробляє травні ферменти, у ній відбувається всмоктування амінокислот, вона є місцем накопичення та зберігання запасних поживних речовин (жирів та каротиноїдів), а також бере участь в екскреції.
У кишці не відбувається ні травлення, ні всмоктування; основна її функція — постачання слизу, що огортає неперетравлені рештки їжі, які викидаються назовні.
Із кишкою пов’язаний дуже своєрідний орган головоногих — чорнильний мішок, в якому утворюється та накопичується чорна рідина. Чорнильний мішок відкривається в кишку поблизу від анального отвору (рис. 106). Він складається з двох частин. Одна з них — чорнильна залоза, клітини якої виробляють гранули пігменту меланіну. Старі клітини поступово руйнуються, їх фарба розчиняється в рідині залози, і утворюється чорнило, яке надходить у другу частину чорнильного мішка —резервуар. При небезпеці тварина викидає через лійку частину вмісту резервуара.
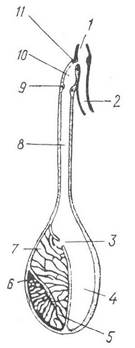
Рис. 106. Схема будови чорнильного мішка Sepia:
1 — анальний отвір; 2 — пряма кишка; 3 — отвір у перетинці; 4 —резервуар; 5 — залозисті складки; 6 — зона їх утворення; 7 — залозистий відділ; 8 — протока чорнильного мішка; 9 — внутрішній сфінктер; 10 — ампула; 11 — зовнішній сфінктер
Основна функція чорнильної рідини —дезорієнтація хижака, який нападає. Викинувши рідину, яка деякий час тримається форми, що нагадує самого молюска, молюск блід- нішає, різко змінює траєкторію руху і зникає, а хижак хапає замість нього його чорнильну копію. Чорнильна рідина подразнює очі хижака і спричиняє тимчасову паралізуючу дію на органи нюху, що перешкоджає хижаку переслідувати здобич.
Чорнильний мішок мають усі кальмари, майже всі каракатиці та більшість безплавцевих восьминогів; немає його у наутилуса, вампіроморф, плавцевих восьминогів та, як виняток, деяких каракатиць. Види, позбавлені чорнильного мішка, мешкають переважно в глибинах океану.
Пігмент головоногих — один із найбільш стійких барвників. Із чорнильних мішків каракатиць здавна виготовляли високоякісну коричневу фарбу — сепію.
Органи виділення головоногих дуже різноманітні та тісно пов’язані з органами кровоносної та дихальної систем (рис. 107). Власне видільна система представлена однією парою (у наутилуса — двома парами) ниркових мішків, які є видозміненими целомодуктами перикардіального відділу целома. Кожна нирка одним кінцем відкривається в перикардій, а другим —у мантійну порожнину. Часто обидві нирки з’єднуються одна з одною поперечною перемичкою або за допомогою непарного мішка, як наприклад у каракатиць. У нирки вдаються ниркові венозні придатки — розростання великих венозних судин, які щільно прилягають до стінок нирок (рис. 108). Через пі придатки протікає венозна кров, з якої нирки вилучають екскрети.
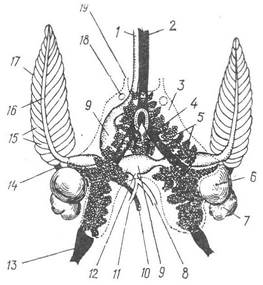
Рис. 107. Видільна, дихальна та центральна частина кровоносної системи каракатиці Sepia:
1 — головна аорта; 2 — головна вена; 3 — нирковий мішок; 4 — порожниста вена; 5 — венозні придатки; б — зяброве серце; 7 — перикардіальна залоза; 8 — анальна артерія; 9 — шлуночок серця; 10 — вена чорнильного мішка; 11 — задня артерія; 12 — черевна аорта; 13 — бічна черевна вена; 14 — зяброва артерія; 15 — зяброві пелюстки; 16 — зяброва вена; 17 — ктенідії; 18 — отвір нирок у перикардій (реноперикардіальний); 19 — зовнішній видільний отвір

Рис. 108. Кровоносна система кальмара Todarodes pacificus (артерії світлі, вени темні):
1, 2 — вени та артерії рук; 3 — глоткова артерія; 4 — артерія переднього краю мантії; 5 — печінкова вена; 6 — кишечна вена; 7 — передня аорта; 8 — зяброва артерія; 9, 10 — печінкова та шлункова артерії; 11 — порожниста артерія; 12 — мантійна вена; 13 — перикардіальна залоза; 14 — зяброве серце; 15 — шлунок; 16, 17 — статеві вена та артерія; 18 — черевна вена; 19 — гонада; 20 — плавцева артерія; 21 — цекум; 22 — мантійна артерія; 23 — задня аорта; 24 — шлуночок серця; 25 — передсердя; 26 — ниркові венозні придатки; 27, 28 — зяброва та головна вени; 29 — печінкова артерія; 30, 31, 32 — вени лійки, мозку та рук відповідно
Крім нирок, видільну функцію виконують також перикардіальні залози, які лежать у відокремлених ділянках перикардіального целома поряд із зябровими серцями (див. рис. 107). З крові зябрового серця речовини, що підлягають видаленню, проникають у порожнину перикардіальної залози, потім у ниркові венозні придатки, де відбувається зворотнє всмоктування солей, амінокислот, цукрів та інших важливих для організму речовин. Отже, основним органом виділення є ниркові придатки; вони виконують також функцію осморегуляції. У ниркових мішках нагромаджується сеча, основним компонентом якої є аміак.
Органами дихання головоногих є ктенідії. У наутилуса їх дві пари, в решти головоногих — одна. Ктенідії розташовані в мантійній порожнині симетрично по боках тулуба. Вони двопірчасті, кожен з них складається із зябрової осі та двох рядів складчастих зябрових пелюсток (див. рис. 107). Загострені вільні кінці ктенідіїв спрямовані вперед. По обидва боки зябрових пелюсток проходять кровоносні судини — приносна та виносна. Епітелій зябрових пелюсток не має війок, і циркуляція води в мантійній порожнині викликається ритмічними скороченнями м’язів мантії.
Кровоносна система досягає в головоногих найбільшої досконалості.
Серце складається з одного шлуночка та двох (підклас Coleoidea) або чотирьох (підклас Nautiloidea) передсердь. Від шлуночка відходять дві аорти — передня та задня (див. рис. 107, 108). Передня, або головна, аорта спрямована вперед до голови і утворює відгалуження (артерії) до переднього відділу кишечника, слинних залоз, печінки; у голові вона розгалужується, утворюючи артерії, що тягнуться вздовж щупалець. Задня, або нутрощева, аорта постачає кров’ю задній відділ кишечника, органи ну трощевого мішка, мускулатуру мантії, статеві органи. Артерії розгалужуються, утворюючи сітку капілярів, з яких беруть початок вени.
Венозні судини розвинені так само добре, як і артеріальні. Венозна система починається венами рук, які впадають у великий кільцевий венозний синус, що збирає венозну кров з голови та рук. Від цього синуса бере початок велика головна вена, яка прямує до нутрощевого мішка і тут ділиться на дві порожнисті вени. Останні вбирають у себе ряд венозних стовбурів від нутрощів. Проходячи біля нирок, порожнисті вени та інші венозні стовбури вдаються туди гроноподібними розширеннями — нирковими придатками; вони слугують для очищення крові від екскретів (див. с. 125).
Порожнисті вени впадають у зяброві серця — скоротливі мішечки, які лежать біля основи зябер. Лише в наутилуса їх немає. Зяброві серця проганяють венозну кров через судини зябер, де вона окислюється і по зябрових венах потрапляє до передсердь, а звідти — до шлуночка. Отже, уся кров, що потрапляє до шлуночка, спочатку проходить через нирки та зябра, і тому в головоногих, на відміну від інших молюсків, серце містить лише артеріальну кров. Кровоносні судини, особливо артерії, мають мускулясті стінки і пульсують, допомагаючи трьом серцям прокачувати кров через капіляри, які особливо розвинені в кінцівках та задній частині мантії.
Кровоносна система головоногих майже замкнена, оскільки в шкірі й м’язах артеріальні капіляри безпосередньо переходять у венозні. В інших місцях між артеріями та венами є синуси. У наутилуса капіляри є тільки в шкірі. Така досконала система кровообігу є одним з факторів, що дає змогу деяким з головоногих досягати велетенських розмірів. Тільки при наявності системи капілярів можливе існування великих тварин, бо лише за цих умов забезпечується живлення та дихання масивних органів.
Нервова система головоногих розвинена по-різному: від дуже примітивної в NautUoidea до найскладнішої і найдосконалішої серед усіх безхребетних у Coleoidea. У наутилуса центральна нервова система складається з трьох коротких нервових дуг —нервових тяжів, які суцільно вкриті нервовими клітинами і не мають диференційованих гангліїв (рис. 109). Усі вони лежать у голові навколо стравоходу. Церебральна дуга огинає стравохід зі спинної сторони, педальна та з’єднана з нею плевровісцеральна — з черевної. Від цих дуг відходять нерви.
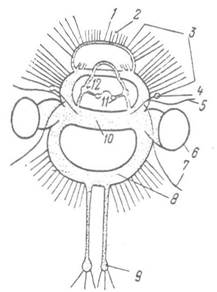
Рис. 109. Центральна нервова система Nautilus:
1 — передпедальне нервове кільце; 2, 3 — нерви відповідно пластинчастого органа та щупалець; 4 — статоцист;
5 — нерви передніх очних щупалець; 6 — око; 7 — нерви задніх очних щупалець; 8 — плевровісцеральний тяж; 9 — черевний ганглій; 10 — церебральний тяж; 11,12 — букальний та лабіальний ганглії
На відміну від наутилуса, центральна нервова система вищих головоногих досягає високої складності будови (рис. 110). Вона дуже концентрована. Церебральні, плевральні, педальні, вісцеральні та парієтальні ганглії тісно згруповані навколо стравоходу й оточені хрящовою головною капсулою. Над стравоходом лежить пара церебральних гангліїв, від яких відходять дуже товсті короткі зорові нерви, що відразу ж розширюються, утворюючи величезні оптичні ганглії. Спереду від церебральних гангліїв розташований невеликий бу- кальний ганглій, зв’язаний з церебральними конективами (у восьминогів він зливається з церебральним). Він іннервує органи глотки та слинні залози. Під стравоходом містяться педальні, плевральні, парієтальні та вісцеральні ганглії. Кожен педальний ганглій чітко поділений на два нервові вузли: брахіальний, або ганглій щупалець, та інфундибулярний, або ганглій лійки. Мозок головоногих, особливо його надстравохідна частина, має складне внутрішнє розчленування, у ньому виділяють окремі зони, які відповідають за певні типи рухових реакцій, складні форми поведінки, пам’ять тощо. Мозок головоногих за об’ємом найбільший серед безхребетних.

Рис. 110. Нервова система Sepia officinalis:
1,2 — нерви та ганглії рук; 3 — верхній букальний ганглій; 4 — нутрощеві нерви; 5 — симпатичний нерв; 6 — задній нерв головної вени; 7 — нерв чорнильного мішка; 8 — його гілки; 9 — ниркові нерви; 10 — серцевий нерв; 11 — нерв нідаментальної золози; 12 — шлунковий ганглій; 13 — шлункові нерви; 14, 15 — нерки сліпого мішка шлунка та прямої кишки; 16, 17 — зяброві ганглій га нерв; 18 — нерв статевої протоки; 19 — зябровий нерв; 20, 21 — мантійні ганглій та нерв; 22 — нерв шийного м’яза; 23 — нюшний нерв; 24, 25 — плевральний та церебральний ганглій; 26 — оптичний ганглій; 27 — очний нерв
Водночас у головоногих за рахунок периферійного нервового плетива з’являються нові, додаткові ганглії, яких немає в інших молюсків. Найбільшими з них є: ганглії щупалець, які залягають уздовж брахіальних нервових стовбурів при основі кожного щупальця; мантійні, або зірчасті, ганглії, що іннервують мантію; букальні ганглії, які іннервують слинні залози та глотку. Крім того, дрібні ганглії розсіяні в товщі мускулатури рук та при основі присосків. Завдяки цьому відрізані руки головоногих зберігають здатність до досить складних та специфічних реакцій на зовнішні подразники.
У головоногих є органи внутрішньої секреції. Це оптичні залози ( у наутилуса їх немає). Оптичні залози — маленькі округлі парні тільця, що лежать на оптичному нерві між оптичними частками та мозком. Вони складаються з секреторних та опорних клітин. Гормони оптичної залози стимулюють розвиток гонад та придаткових статевих залоз, управляють формуванням сперматофорів, визначають поведінку тварин, пов’язану з розмноженням та турботою про нащадків. Активність оптичних залоз контролюють певні центри мозку. Ці залози також беруть участь у захисті організму від чужорідних білків.
Нейросекреторні клітини є в різних місцях центральної та периферійної нервової системи. Нейрогормони виділяються в кров і впливають на вегетативні функції організму (регулюють роботу серця, тиск крові тощо).
Органи чуття в головоногих розвинені дуже добре. У них є статоцисти, пара очей, позаочні фоторецептори, нюшні ямки, субрадулярний орган, жому приписують функцію органа смаку, а також окремі чутливі клітини на присосках рук та шкірі.
Пара статоцистів міститься в голові. Це зближені між собою пухирці, вкриті окремими хрящовими капсулами, пов’язаними з головною хрящовою капсулою. Внутрішня поверхня статоциста має опуклості та горби, що вдаються в порожнину органа (рис. 111). У певних місцях горбків містяться чутливі нервові клітини. Статоліт великий, неправильної форми, він складається з органічної речовини і частково з вуглекислого кальцію. Видалення статоцистів викликає втрату молюском здатності до орієнтації в просторі.

Рис. 111. Будова статоцистів Sepia officinalis (зріз через голову):
1 — підстравохідна частина мозку; 2 — стравохід; 3 — статоліт; 4 — горбки внутрішньої поверхні статоциста; 5 — слуховий гребінець; 6 — перегородка між статоцистами; 7 — капсула статоциста
Будова очей Nautiloidea та Coleoidea дуже розрізняється (рис. 112). Очі наутилуса побудовані за типом очного пухиря; такі очі є у багатьох безхребетних (деяких медуз, кільчаків, двостулкових та черевоногих молюсків).
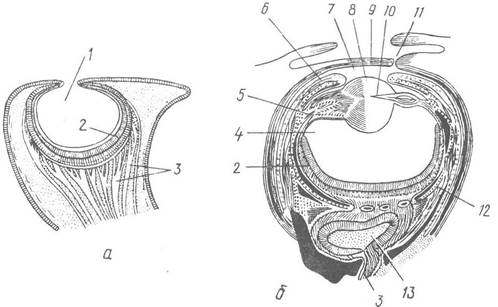
Рис. 112. Очі головоногих:
а, б — розріз ока Nautilus та Sepia officinalis; 1 — порожнина очної ямки, яка сполучається із зовнішнім середовищем; 2 — сітківка; 3 — зоровий нерв; 4 — склоподібне тіло; 5 — війковий м’яз; 6 — райдужка; 7— передня камера ока; 8 — кришталик; 9 — рогівка; 10 — епітеліальне тіло; 11 — зовнішній отвір камери ока; 12 — склера (хрящова оболонка ока); 13 — оптичний ганглій
Очі Coleoidea відрізняються складністю та досконалістю будови і нагадують очі хребетних. Вони досягають великих розмірів, займаючи у деяких видів більшу частину голови. Зовні око оточене хрящовою капсулою. Основу ока складає очний пухир. Його дно та бічні стінки утворюють сітківку, або ретину. Зовнішня стінка щільно прилягає до покривів, утворюючи внутрішнє епітеліальне тіло, а прилеглий до неї покривний епітелій утворює зовнішнє епітеліальне тіло. Епітеліальне тіло виділяє кришталик, причому зовнішнє тіло виділяє зовнішню його половину, а внутрішнє —внутрішню. Порожнина пухиря заповнена склоподібним тілом. Над передньою стінкою очного пухиря наростає кільцеподібна складка шкіри у вигляді купола — райдужка; її краї не змикаються в центрі, залишаючи отвір над кришталиком — зіницю. Над райдужкою утворюється друга, зовнішня складка шкіри, яка покриває зрачок та кришталик і перетворюється на передню прозору стінку ока — рогівку. Проте у більшості головоногих вона не повністю змикається над оком, зберігаючи маленький ексцентричний отвір, через який передня камера ока сполучається із зовнішнім середовищем. Крім того, у каракатиць та деяких восьминогів рогівку прикривають ще шкірні повіки. Сітківка ока складається з дуже довгих (іноді до 0,5 мм) клітин. Зорові клітини правильно чергуються з опорними, які містять темний пігмент. Сукупність нервових відростків, що відходять від зорових клітин, утворює товстий зоровий нерв, який веде в дуже великий зоровий ганглій. Велика кількість зорових клітин сітківки (у кальмара Loligo близько 165 тис.) свідчить про досконалість зору.
Очі головоногих здатні до акомодації, яка здійснюється не зміною кривизни кришталика (як у людини), а його наближенням або віддаленням від сітківки; для цього служить особливий війковий м’яз, прикріплений до екватора кришталика. Крім того, у райдужці є м’язи, які розширюють або звужують зіницю залежно від інтенсивності освітлення.
Позаочні фоторецептори — загадкові органи головоногих. Вони є в усіх головоногих, крім наутилуса. Це скупчення пухирців, які містять світлочутливі клітини та пов’язані з нервовою системою. Вони можуть бути розташовані в різних частинах тіла. Наприклад, у восьминогів вони містяться на задній стороні зірчастого ганглію, у каракатиць — в голові на оптичних нервах, а в кальмара-вампіра — у м’язах спинної сторони мантії. Вважають, що вони сприймають світло, яке проходить крізь стінку тіла, і дають тварині уявлення про
рівень освітлення в оточуючому середовищі; залежно від його інтенсивності головоногі регулюють силу власного свічення. Можливо, що за допомогою цих органів головоногі сприймають біолюмінісценцію інших тварин. Крім того, головоногі сприймають світло також за допомогою численних світлочутливих клітин, розсіяних у їхній шкірі.
Хеморецепторів у головоногих є кілька типів. Органи нюху — це пара нюшних папіл (у кальмарів, каракатиць та пелагічних восьминогів) або нюшних ямок (у донних восьминогів), які містяться по боках голови між очима та мантійним отвором. У наутилуса органами нюху є ринофори — пара невеличких конічних горбків з вузькою порою, які містяться під очима.
Смакові рецептори розташовані переважно на обідках присосків рук та на губі. Це різноманітні війчасті рецепторні клітини. Кількість їх величезна: до кількох сотень на 1 мм2. Головоногі мають дуже тонкий смак; їх чутливість до деяких хімічних речовин на 2—3 порядки вища, ніж у людини. На присосках містяться також численні механорецептори, особливо дотичні, які реагують на стиснення, розтягання та згинання. Тісна близькість смакових та дотичних рецепторів дає підставу говорити про наявність у головоногих особливого хемотактильного — «смакодотичного» — чуття. Особливо воно характерне для донних та глибоководних видів. У донних восьминогів добре розвинена хемотактильна пам’ять, у тому числі здатність знаходити дорогу до своєї домівки. Крім того, у порожнині глотки є так званий субрадулярний орган, який містить чутливі клітини і якому приписують функцію органа смаку. У наутилуса органами дотику й смаку є щупальця; на відміну від інших головоногих наутилус має пару осфрадіїв, які лежать в мантійній порожнині і використовуються для визначення хімічних властивостей води, що надходить до неї.
У головоногих є безліч внутрішніх та зовнішніх пропріоцепторів у шкірі, м’язах, товщі тканин присосків, поверхні зябер тощо. Вони надають тварині інформацію про відносне положення частин її тіла, роботу мускулатури.
Головоногим притаманні складні форми поведінки. Передусім вони виявляються в реакціях при нападі на здобич та втечі від ворогів. Не менш складна поведінка супроводить запліднення, відкладання та охорону (у восьминогів) самицями яєць. Головоногі, особливо донні восьминоги та каракатиці, мають пам’ять, досить легко навчаються. З іншого боку, наутилусам, пелагічним восьминогам та глибоководним океанічним кальмарам ці здібності притаманні значно меншою мірою.
Усі головоногі роздільностатеві тварини з чітко вираженим статевим диморфізмом. Самці відрізняються від самиць тим, що одне з їх щупалець перетворене на копулятивний орган (гектокотильоване щупальце). У деяких головоногих, наприклад у Argonauta, самець набагато менший за самицю.
Статева залоза непарна і міститься в статевій ділянці целома, у задній частині тіла. Статеві клітини потрапляють до порожнини целома, а звідти виходять через вивідні протоки. У Nautilus, вампіроморф та плавцевих восьминогів статеві протоки парні, в інших головоногих зберігається здебільшого лише ліва протока (рис. 113). Крім того, незалежно від статевого отвору, але поблизу від нього в мантійну порожнину відкриваються вивідні канали двох парних та однієї непарної нідаментаяьних заюз, секрет яких утворює зовнішні оболонки яєць. У багатьох головоногих є сім’яприймачі — пара ямок, які містяться на ротовому конусі.
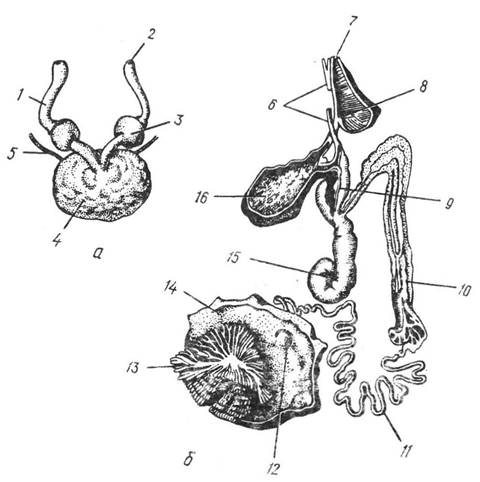
Рис. 113. Статева система восьминога Octopus dofleini:
а — жіноча; б — чоловіча; / — яйцепровід; 2 — жіночий статевий отвір; 3 — яйцепровідна залоза; 4 — яєчник (у целомічному мішку); 5 — зв’язка, яка підтримує яєчник; 6 — перерізаний м’яз; 7 — чоловічий статевий отвір; 8 — розширення статевого каналу; 9 — з’єднання сім’яного пухирця і простатичної залози зі сперматофорним мішечком; 10 — сім’яний пухирець; 11 — сім’япровід; 12 — отвір сім’япроводу; 13 — сім’яник (всередині целомічного мішка); 14 — спяки целомічного мішка; 15 — простатична залоза: 16 — сперматофорний мішок
Чоловіча статева протока здебільшого непарна і має складнішу будову. Сім’япровід відкривається в статевий це лом лійкою; на початку він утворює розширення — сперматофорний орган, який має складну будову. До його складу входять залозистий сім’яний пухирець, простатична залоза, сліпий мішок та інші утвори. Далі сім’япровід продовжується широким сперматофорним мішком, який переходить у копулятивний орган (пеніс), що закінчується статевим отвором.
Запліднення самиць відбувається за допомогою сперматофорів. Стінки сім’яного пухирця та простатичної залози виділяють речовину, яка оточує сперму, утворюючи сперматофор складної будови (рис. 114). Він має видовжену форму, його стінки складаються з хітиноїдної речовини. Сперматофори накопичуються в сперматофорному мішку. Вони мають особливі пристосування для своєчасного звільнення сперми, яка в них знаходиться. Під дією морської води зрілі сперматофори лопаються і викидають сперму, яка міститься в них.

Рис. 114. Сперматофор Octopus doileini
Роль копулятивного органа виконує одна із рук самця, яка більш-менш відрізняється від інших і називається гектокотилем (рис. 115). Цією рукою самець підхоплює сперматофори, які виходять через лійку назовні, і переносить їх у мантійну порожнину самиці (у восьминогів) або прикріплює до сім’яприймачів самиці, які містяться на її ротовому конусі (у Nautilus, Sepia, Sepiola, Loligo та деяких інших кальмарів).

Рис. 115. Дві стадії розвитку гектокотиля Argonauta argo:
а — самець зі сперматофорним мішком (1); б — зі сформованим гектокотилем (2)
Надзвичайне пристосування до запліднення є в дрібних пелагічних восьминогів — аргонавтів, тремоктопусів, оцитос. Дуже великий гектокотиль у самців розвивається в особливому шкірястому мішку, де він спочатку згорнутий у спіраль. Коли щупальце повністю сформувалося, мішок розривається, і воно розправляється (рис. 115), а його порожнина заповнюється сперматофорами. Щупальце відривається від тіла самця і відпливає на пошуки самиці свого виду. Знайшовши самицю, гектокотиль заповзає в її мантійну порожнину. Там сперматофори, які він приносить, лопаються, і сперматозоїди запліднюють яйця. Втрачений гектокотиль згодом регенерує.
Спочатку дослідники, знаходячи гектокотилі в мантійній порожнині самиць, вважали їх паразитами, і Кюв’є дав їм родову назву Hectocotilus.
Запліднення яєць проходить здебільшого під час їх відкладання або в мантійній порожнині самиці, або коли вони викидаються через лійку та проходять повз рот, де містяться сім’яприймачі зі спермою. Лише в аргонавтів яйця запліднюються ще в яйцепроводі. У деяких видів, наприклад восьминога Ocythoe, яйця затримуються в яйцепроводі до виходу з них молоді, тобто має місце живородіння. Здебільшого ж яйця, оточені оболонками, виводяться назовні і прикріплюються поодинці чи групами до різних підводних предметів або містяться всередині драглистих мішків, які вільно плавають у воді.
Яйця головоногих великі і містять багато жовтка. Через це ембріональний розвиток головоногих набуває особливостей, які відрізняють його від розвитку інших молюсків. Сліди спірального дробіння яйця зникають, і воно стає дискоїдальним.
Жовток заповнює майже все яйце, лише на анімальному полюсі лежить дископодібне потовщення цитоплазми, яке містить ядро. Дробіння охоплює лише анімальний полюс: тут утворюється спочатку одношаровий, а пізніше двошаровий зародковий диск (рис. 116, а). Для переробки жовтка виникає особливий провізорний орган — жовтковий мішок, стінки якого складаються з ектодерми та жовткової ентодерми з кровоносними лакунами та м’язовими елементами між ними. Поживні речовини із жовткового мішка транспортуються до зародка.

Рис. 116. Ембріональний розвиток кальмара Loligo vulgaris:
а — дробіння яйцеклітини; б — утворення зачатків органів; в — завершення формуван ня молюска; 1 — зародковий диск; 2 — жовток; 3 — черепашкова залоза; 4 — зачаток лійки; 5 — рот; 6 — зачатки рук; 7 — жовтковий мішок; 8, 9 — зачатки очей та плавців; 10 — мантія
З двошарового зародкового диска формується тіло зародка. На його анімальному полюсі з’являється ектодермальне потовщення з невеличким впинанням посередині — зачатком черепашкової залози; край цього потовщення стає мантією. Пізніше у всіх колеоідей черепашкова залоза перетворюється на замкнений мішечок, який зовні огортає мантія.
Так виникає внутрішня черепашка, а в Argonauta черепашкова залоза зовсім зникає. На спинній стороні зародка утворюються зачатки очей у вигляді потовщення ектодерми, а між ними — рот. На черевній стороні закладаються зябра, зачаток лійки, статоцисти, а на межі власне зародка і жовткового мішка — руки. Зародок розташований так, шо головним кінцем він обернений до жовткового мішка і ніби охоплює його зачатками рук (рис.116, б, в). Пізніше зародок збільшується, а жовтковий мішок зменшується і втягується всередину зародка.
З яйцевих оболонок виходить маленький, майже повністю сформований молюск (Octopus, Nautilus, Sepia). Проте у багатьох видів молодь суттєво відрізняється від дорослих молюсків формою тіла та наявністю личинкових органів, яких немає в дорослих, тому таких молодих особин називають личинками. Іноді личинок вже відомих видів описували як самостійні види або навіть роди. Наприклад, личинки кальмарів родини хіротеутид (доратопсис) мають дуже довгу шию та «морду», личинки кальмарів родини кранхіїд — стебельчасті очі, а личинки пекельного вампіра — дві пари плавців, тоді як дорослі — одну пару (див. рис. 119). Часто личинки ведуть планктонний спосіб життя і трапляються на менших глибинах, ніж дорослі.
Головоногі мають високорозвинену здатність до регенерації пошкоджених або втрачених частин тіла. Рани на їх тілі заживають дуже швидко. Втрачені (наприклад, відкушені хижаком) руки та щупальця швидко повністю відновлюються. У багатьох видів океанічних кальмарів та восьминогів спостерігається автотомія — довільне відкидання кінцівок. Здебільшого це відбувається при небезпеці. У аргонавтів та деяких інших груп гектокотилізоване щупальце із сперматофором відривається та заповзає у мантійну порожнину самиці.
Способи руху головоногих дуже різноманітні, відповідно різні й біологічні механізми, що забезпечують той чи інший рух. Найдосконалішим є реактивний рух, який забезпечується роботою мантії та лійки. Мантійна порожнина головоногих дуже велика. У кальмарів її об’єм становить біля половини об’єму тіла. Набираючи в неї воду через мантійну щілину, молюск із силою виштовхує її потім через лійку. Щоб вода при цьому не протікала назад через щілину, він герметично замикає її за допомогою застібок-«кнопок», про які вже згадувалося.
Коли молюск скорочує мускулатуру черевної стінки мантії, сильний струмінь води викидається з лійки; реактивна сила, яка виникає при цьому, штовхає тіло тварини в протилежний бік. Реактивні поштовхи повторюються з великою частотою, що забезпечує високу швидкість руху.
Лійка в нормальному положенні напрямлена отвором уперед, отже, при реактивному русі молюск рухається вперед заднім кінцем. Проте мускулатура лійки дає змогу їй повертатися отвором у різні боки, навіть на 180 °, спрямовуючи його назад, що забезпечує тварині можливість рухатись у різних напрямах, у тому числі й головою вперед.
Деякі невеликі за розмірами кальмари розвивають таку швидкість, що можуть вистрибувати з води і пролітати над поверхнею моря по 50—60 м. Часто вони залітають на верхні палуби океанських лайнерів, які знаходяться на висоті 5— 8 м над рівнем моря. Планеруючий політ підтримується за допомогою широких плавців. У такий спосіб кальмари рятуються від хижих риб та дельфінів у поверхневих шарах води.
Своєрідну будову має реактивний двигун наутилуса. Його мантія майже позбавлена м’язів, і функцію ємкості, в яку набирається вода, виконує не мантійна порожнина, а велика, добре розвинена лійка. Вона складається з двох трикутних м’язистих лопатей, які при русі згортаються в трубку, налягаючи одна на одну. При скороченні мускулястих стінок із лійки викидається струмінь води, який створює реактивний рух.
Реактивний рух створюється не тільки роботою комплексу мантія—лійка, але й руками. У деяких головоногих, особливо глибоководних пелагічних восьминогів, усі руки з’єднані шкірною перетинкою, яка утворює «парасольку». Її краї розростаються так широко, шо досягають кінчиків рук. Парасолька восьминогів нагадує дзвш медуз, і діє вона за тим же принципом: при розкритті дзвона вода заповнює простір між руками, а при його скороченні виштовхується назовні, і тварина рухається в протилежний бік (заднім кінцем уперед). Порівняно з реактивним двигуном кальмарів та каракатиць цей механізм менш досконалий і не дає змогу восьминогам швидко рухатись.
Реактивний рух —не єдиний спосіб пересування головоногих. Вони плавають також за допомогою плавців, які є в кальмарів, каракатиць, вампіроморф та плаваючих восьминогів. Решта восьминогів живе постійно на дні і плавців не має. Плавці забезпечують повільне плавання та ширяння тварин у воді. Взагалі, нектонні кальмари та каракатиці мають щільне мускулясте тіло, яке важче за воду, і щоб не потонути, вони повинні весь час рухатися.
Основним способом руху донних восьминогів є повзання по дну за допомогою рук. Вони навіть можуть крокувати по дну, спираючись на кінчики рук.
Поряд із активно-рухливими тваринами серед головоногих є також планктонні організми. Одні з них мають драглисте тіло і більше схожі на медуз, ніж на головоногих; інші мають тонку, майже прозору мантію, позбавлену м’язів. Ці тварини живуть, як правило, на глибині 100 і більше метрів і ширяють у воді. Вони мають нейтральну плавучість, тобто їхня питома маса наближається до питомої маси води.
У головоногих існує два способи досягнення нейтральної плавучості: за допомогою черепашки, камери якої можуть заповнюватися газом (Nautilus, Sepia, Spirula), та шляхом зменшення питомої маси тканин тіла (глибоководні кальмари та деякі інші). Перший механізм нейтральної плавучості дуже ефективно діє у наутилуса. Як уже зазначалося, камери його черепашки пронизує сифон —відросток нутрощевого мішка, в який заходять целом та кровоносні судини. Стінка сифона — напівпроникна мембрана, яка пропускає лише одновалентні іони та гази.
У положенні нейтральної плавучості камери черепашки або повністю заповнені газом, або води в них дуже мало. Якщо необхідно зануритися на глибину, тобто загрузити черепашку, у камери додається вода з крові, а для того, щоб сплисти, з води вилучаються іони Na та Сl, вода опріснюється, і черепашка полегшується. Отже, сифон —це спеціалізований орган для регуляції плавучості.
У планктонних кальмарів нейтральна плавучість досягається іншим способом. Здебільшого в товщі тканин їх мантії, голови та рук містяться численні мікроскопічні вакуолі, заповнені розчином хлориду амонію (NH4Cl). Утворюється губчаста тканина, яка замінює собою мускульну. Розчин хлориду амонію ізотонічний щодо морської води, тому питома маса планктонних кальмарів наближається до питомої маси води. Таких кальмарів називають аміачними; до них належать переважно глибоководні види. Найдосконаліше пристосування такого типу мають океанічні кальмари родини Cranchlidae — «кальмари-батискафи». У них розчин хлориду амонію заповнює целом, його об’єм може досягати 2/3об’єму мантійної порожнини.
З відомих 25 родин кальмарів нейтральна плавучість властива представникам 12 родин. Аміачні кальмари —переважно дрібні тварини, але серед них є й крупні, наприклад велетенські кальмари Architeuthis та Mesonychoteuthis.
Головоногі відіграють дуже важливу роль у житті океану. Будучи хижаками, вони поїдають величезну кількість ракоподібних, молюсків, риб та інших організмів і, у свою чергу, самі є їжею для багатьох морських хребетних — риб, птахів, ластоногих та китів.
Людина здавна використовувала головоногих в їжу, а в останні десятиріччя промисел головоногих різко посилився, що пов’язано з виснаженням рибних ресурсів та необхідністю пошуків додаткових джерел білкової їжі. За поживними якостями головоногі перевершують інших молюсків та навіть деяких риб. В Японії це один із найпопулярніших продуктів харчування. У наш час існує промисловий вилов близько 30 видів головоногих. Це передусім кальмари омастрефіди, лолігініди, деякі каракатиці та звичайні восьминоги. Із гарних черепашок наутилусів роблять прикраси; з чорнильної рідини сепій виготовляють фарбу та чорнило.
Головоногих використовують також у медицині та парфюмерії. Новітніми дослідженнями доведено, що головоногі є джерелом важливих та перспективних біологічно активних препаратів (протишокових, обезболюючих, антипаразитарних речовин, сильнодіючих токсинів тощо).
Головоногі викликають інтерес вчених як модельні об’єкти досліджень: для нейрофізіологів кальмари — один із важливих об’єктів для дослідження проведення нервових імпульсів; психологи цікавляться головоногими як безхребетними з високорозвиненою психікою («примати моря»).
Сучасні Cephalopoda поділяються на два підкласи: Наутилоідеї (Nautiloidea) та Колеоідеї (Coleoidea).