ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 3 - Г.Й. Щербак - 1997
ОГЛЯД БЕЗХРЕБЕТНИХ ТВАРИН ПО ЕРАХ ТА ПЕРІОДАХ
Тварини з’явились на Землі в протерозойську еру, однак для розуміння передумов їх виникнення слід мати певну уяву і про більш ранні етапи розвитку життя на нашій планеті.
Архейська ера. На початку архею утворились такі оболонки Землі, як літосфера, гідросфера та атмосфера. Залишки перших живих організмів мають вік близько 3,5 млрд років. Це мікроскопічні кулясті, паличкоподібні чи зірчасті форми, що, безумовно, належать до бактерій, та строматоліти. Останні — це вапнякові оболонки колоній ціанобактерій, або синьозелених «водоростей»; подібні види існують і досі. Отже, в архейську еру в морях існували екосистеми, що складалися з автотрофних та гетеротрофних прокаріотичних організмів. Завдяки їх діяльності утворились поклади залізної руди, фосфоритів, графіту тощо. Фотосинтезуюча діяльність ціанобактерій привела до того, що наприкінці ери вміст кисню в повітрі мало відрізнявся від сучасного. Це дало змогу для розвитку енергетично економного аеробного дихання, що відкрило дорогу до інтенсифікації обміну речовин і подальшої еволюції живих істот.
Протерозойська ера. У цю еру виникають еукаріоти, а пізніше — багатоклітинні організми, у тому числі й тварини з променевою чи двобічною симетрією. Життя існувало тільки в морях; постійних прісних водойм тоді ще не було. Суходіл являв собою мертву пустелю. Залишки перших еукаріот, подібних до сучасних джгутикових, відомі починаючи з середини ери; у викопних мулах та стінках строматолітів знайдено ходи, які, безумовно, були зроблені якимись багатоклітинними тваринами. Однак багата фауна безхребетних розвивається лише наприкінці протерозою — у венді.
Вендський період був досить теплим; існував єдиний суперконтинент, оточений мілководними морями, де переважно й мешкали тварини. Фауна венду характеризується досить великими (до метра завдовжки чи в діаметрі) за розмірами тваринами без будь-яких скелетних утворів або черепашок. Відсутність багатоклітинних некрофагів призвела до того, що залишки цих організмів добре збереглись у скам’янілому мулі у вигляді відбитків. З цього періоду відомо багато залишків особливих багатоклітинних водоростей (вендотеніди), ймовірно, зелених, які складали багату харчову базу для рослиноїдних тварин.
Близько 2/3 фауни венду складали різноманітні кишковопорожнинні з класів Scyphozoa та Hydrozoa, а також відомі лише з цього періоду представники класів Cyclozoa, Inordozoa та Trilobozoa, які описані в першій книзі трьохтомного видання підручника.
Повзаючих, плаваючих чи сидячих вендських безхребетних з двобічною симетрією за браком даних про їх будову важко віднести до відомих систематичних груп: це так звані проблематики венду. Серед них були несегментовані й сегментовані форми; у деяких з останніх був добре розвинений головний відділ та нечленисті кінцівки. На межі венду та кембрію єдиний суперконтинент розпався на окремі платформи, що призвело до майже повної загибелі вендської біоти. У нових умовах морські мілководдя стали осередками розвитку наступної, кембрійської, фауни.
Палеозойська ера. Це час виникнення всіх типів та класів безхребетних; життя опановує прісні водойми та суходіл. Виняткове значення в історії розвитку безхребетних має кембрійський період.
Для безхребетних кембрійської фауни надзвичайно характерне виникнення захисних структур (черепашок) або захисно-рухових систем (скелетів). На відміну від сучасних видів, скелет у яких переважно вапняковий, у кембрії були поширені, поряд з вапняковими, і фосфатні скелети (наприклад, у кембрійських Brachiopoda; з їхніх залишків складаються поклади фосфоритів в Естонії тощо). Залишки кембрійських тварин знаходять в усіх частинах світу, однак найкраще збережені скам’янілі тварини відомі зі сланців Берджис у Канаді; американські вчені зробили багато достовірних реконструкцій цих організмів; деякі з них будуть розглянуті далі.
Єдиний вендський суперконтинент у кембрії розпався на великий материк південної півкулі — Гондвану та кілька північних материків — Європейський, Сибірський, Китайський та Північноамериканський. До складу Гондвани входили Південна Америка, Африка, Аравія, Південно-Східна Азія, Австралія та Антарктида.
У кембрії життя концентрувалось на мілководдях, де температура води була в межах 25—35°С. Багата флора придонних зелених та червоних водоростей і добре розвинений фітопланктон складали харчову базу тварин. Вже з раннього кембрію відомі найпростіші — форамініфери та радіолярії. Кембрійські форамініфери мали однокамерну аглютиновану дископодібну черепашку (рід Lukatiella), а радіолярії —простий сферичний скелет з двоокису кремнію. Цікаво, що кембрійські радіолярії, на відміну від сучасних, мешкали лише на мілководді — один з доказів того, що в цей період глибини океану ще не були освоєні живими організмами. З губок домінували археоціати, які поряд зі строматолітами утворювали рифи; відомі також численні знахідки спікул Demospongiae. У ранньому кембрії з’являються перші рецептакуліти, що належали до особливого класу Radiocyatha.
На відміну від венду, кишковопорожнинні в кембрії нечисленні. Це група поліпів Hydroconozoa, залишки яких являють собою невеликі (кілька міліметрів) порожнисті конуси, та окремі відбитки сифонофор та сцифомедуз, а також пірамідальні скелети сидячих сцифоїдних поліпів з окремого підкласу Conulata (рис. 239,а). Нещодавно в СІЛА та Росії (у Східних Саянах) знайдено залишки кембрійських видів ряду Hydrida.
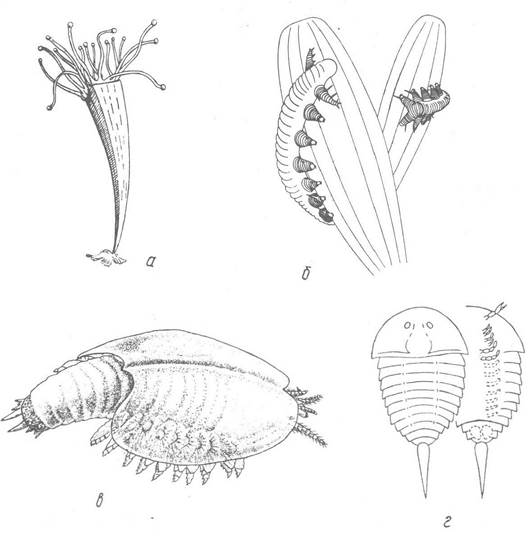
Рис. 239. Реконструкції представників кембрійської фауни:
а — Archaeoconularia (клас Scyphozoa, підклас Conulata); б — Aysheaia (тип Onychophora); в — Canadaspis (клас Malacostraca); г — Aglaspella (клас Xyphosura)
Приапуліди відомі починаючи з середнього кембріно; вони нічим не відрізнялись від сучасних, сягали значної видової різноманітності та, ймовірно, займали ту саму адаптаційну зону, що й зараз — бентосні риючі хижаки. Добре відомі трубки сидячих поліхет та кілька представників Errantia, наприклад Canadia. З середнього кембрію відомий єдиний викопний представник Onychophora — Aysheaia pedunculata (рис. 239, б), який, ймовірно, живився губками; яким чином і коли оніхофори вийшли на суходіл, невідомо.
У морях мешкає багато членистоногих. Це, по-перше, різноманітні зябродишні. Усі вони жили на мілководді морів: щитні (Notostraca), вусоногі (Cirripedia), черепашкові рачки (Ostracoda) та один представник Malacostraca, що належить до викопного ряду Archaeostraca — Canadaspis (рис. 239, в). З раннього кембрію відомо близько 50 видів трилобітоподібних; на кінець періоду вони досягли значної видової різноманітності, освоїли різні адаптаційні зони та стали однією з домінуючих груп. З хеліцерових знайдено мечохвостів, які належали до особливого кембрійського ряду Aglaspidida (рис. 239, г).
Багато форм членистоногих, знайдених переважно в сланцях Берджис (Канада), не належить до жодного з відомих класів. Розглянемо найцікавіші з них.
Marrella splendens (рис. 240, а) — бентосна тварина до 2 см завдовжки, сегментацією тіла нагадувала трилобітів, але на відміну від останніх мала на голові дві пари напрямлених назад загострених відростків; сегменти її тулуба не мали характерних для трилобітів бічних виростів. Канадські вчені запропонували виділити для цього виду окремий клас — Marrellomorpha, близький до Trilobitomorpha.
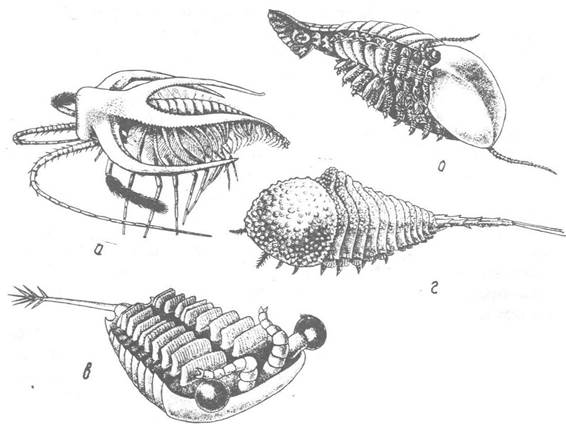
Рис. 240. Реконструкції кембрійських членистоногих зі сланців Берджис (Канада):
а — Маrrеlіа; б — Sidneyla; в — Sarotrocercus; г — Habelia
Sidneyia (рис. 240, б) мала великі розміри (до 20 см); подібно до річкового рака повзала по дну й плавала. її широке плоске тіло складалося з протоцефалона, тулуба та хвостової частини. З боків протоцефалона розташовувались парні очі та багаточленикові антени; кінцівки тулуба були одногіллястими з жуйними відростками; п’ять пар задніх кінцівок були коротші за передні і мали зяброві вирости. Хвостова частина складалась із трьох сегментів без кінцівок та лопатеподібного тельсона.
Sarotrocercus (рис. 240, в) — тварина, що плавала на спині. Тіло її складалося з протоцефалона, тулуба та шилоподібного тельсона з кількома голками на кінці; тулубні кінцівки перетворені на гребені пластинки.
Habelia (рис. 240, г) — бентосна тварина з широким, опуклим зверху тілом, що складалося з голови, вкритого товстим шипуватим панцирем тулуба та вузької хвостової частини з шипами.
З початку кембрію відомі такі групи молюсків, як моно- плакофори, гастроподи, двостулкові (рід FordiUa), стенотекоїди, ксеноконхії, хіоліти. Головоногі з’являються в середньому кембрії; вони мали конічну черепашку та належали до особливого ряду наутилоідей — Plectronoceratida. Brachiopoda (як замкові, так і беззамкові) відомі починаючи з раннього кембрію, а наприкінці періоду стають рифоутворювачами та домінують у морях разом із трилобітами. З кембрію відомі також і фороніди. У кембрійських відкладах знайдено трубки якихось бентосних тварин, яких було названо Sabelliditida. Тривалий час їх вважали близькими до сидячих Polychaeta, однак сучасні дослідження показали, що насправді це трубки Pogonophora. У кембрії були досить поширені як сидячі, так і повзаючі Echinodermata, що належали до восьми класів. Із сидячих (підтип Crinozoa) відомі лише представники вимерлого ряду Eocrinoidea, з повзаючих — Edrioasteroidea та Ноlothuroidea, а також представники ще деяких маловідомих вимерлих наприкінці періоду класів. У середньому кембрії з’являються й представники Hemichordata з класу Pterobran- chia, представлені бентосними та псевдопланктонними колоніями.
Велика кількість кембрійських видів належить до проблематиків, наприклад хіоліти, яких все ж більшість дослідників вважає особливою групою молюсків. Нижче наводяться найцікавіші з проблематиків, які не мають аналогів ні серед викопних, ні серед сучасних (рецентних) таксонів.
Hallucigenia (рис. 241, a) — бентосна тварина 3—5 см завдовжки з м’яким нечленистим тілом. Голова сферична, без будь-яких придатків чи отворів; тулуб циліндричний з сімома парами нечленистих кікцівок-«ходуль» та такою самою кількістю м’яких трубок, напрямлених догори; хвостова частина у вигляді видовженої трубки з отвором.

Рис. 241. Реконструкції тварин-проблематиків кембрію:
а — Hallucigenia; б — Wiwaxia; в — Anomalocaris; г — Nectocaris; д — Odontogriphus; е — Dinomischus;
є — Amiskwia; ж — Opabinia
Wiwaxia —невеликі (5—6 см завдовжки) бентосні повзаючі тварини овальної форми з пласкою нижньою та опуклою верхньою сторонами. Нечленисте тіло зверху вкрите листоподібними лусочками, серед яких виділяються два поздовжніх ряди ножеподібних жорстких утворів, напрямлених догори (органи захисту).
Anomalacaris — плаваючі тварини; описано два види цього роду, які досягали значних розмірів (70—100 см завдовжки). Тіло їхнє складалося з голови й тулуба. Голова мала вигляд суцільної капсули з парою великих очей на коротеньких стебельцях та парою багаточленикових щупалець, кожен членик яких мав пару видовжених виростів із гострими шипами. Вважають, що щупальцями тварина захоплювала їжу і спрямовувала її у рот, оточений диском, на якому розташовувались кілька рядів хітинових зубців. Видовжений плескатий тулуб мав слабо виражену сегментацію та ніс кілька пар бічних нечленистих м’яких лопатей, які утворювали гребний апарат.
Nectocaris —тварина, тіло якої складалося з головного та тулубного відділів без кінцівок. Голова з видовженими бічними лопатями мала одну-дві пари коротеньких загострених на кінці щупалець та пару очей. Сплющений з боків видовжений тулуб складався приблизно з сорока сегментів, мав довгий спинний та короткий черевний плавці.
Odontogriphus — тварина, яка мала близько 8 см завдовжки і вигляд широкого сплющеного в дорзовентральному напрямку черва. Тіло складалось із напівкруглої голови та членистого (близько ЗО сегментів) тулуба, позбавленого будь-яких кінцівок. Знизу голови була розташована пара сенсорних органів у вигляді конічних заглибин та рот, оточений V-подібним рядом конічних виростів.
Dinomischus — сидячі тварини завдовжки 4—5 см. Тіло складалося з чашечки та стебельця. Диск чашечки був оточений нерозгалуженими пелюсікоподібними утворами, які щільно з’єднувались між собою бічними краями. Посередині диску містився рот, трохи збоку від нього —анальний отвір.
Amiskwia була плаваючою твариною завдовжки близько 4 см, зовні дещо подібною до Chaetognatha, але без характерних для них щелеп та капора. Нечленисте, з м’якими тонкими покривами тіло складалося з голови та тулуба. На голові була розташована пара щупалець; тулуб мав пару бічних плавців та широкий горизонтальний хвостовий.
Opabinia — плаваюча хижа тварина. Вона мала червоподібне тіло, яке складалося з суцільної голови, багатосегментного тулуба та заднього відділу з трьох сегментів. На голові були розташовані п’ять очей та довгий, напрямлений уперед хоботок, який мав вигляд гнучкої циліндричної трубки, а на кінці були дві лопаті з довгими голками на внутрішніх
поверхнях. Можливо, тварина хоботком захоплювала їжу, потім він згинався і подавав її до рота. Сегменти тулуба мали бічні пластинчасті вирости із зябрами, а сегменти хвостового відділу — по парі округлих пластинок, напрямлених вгору і вбік; ймовірно, цей відділ грав роль руля при плаванні. Непочленовані кінцівки та наявність гнучкого хобота, який, мабуть, мав гідравлічний принцип дії, свідчать про відсутність екзоскелета в цієї тварини.
Наприкінці кембрійського періоду відбуваються зміни обрисів материків, опускання морського дна тощо, що зумовило часткову руйнацію екосистем та вимирання ряду груп: поліпів Hydroconozoa, мечохвостів ряду Aglaspidida, стенотекоїдів; повністю зникають проблематики кембрію, майже цілком — археоціати.
Ордовицький період характеризується тим, що живі організми освоїли всі глибини Світового океану та прісні водойми. У геологічному відношенні це був період переважання морів над суходолом; він був такий самий теплий, як і кембрій. Значно збільшується чисельність та різноманітність форамініфер і радіолярій; вони стають породоутворювачами. Дуже поширені звичайні й скляні губки. Досягають розквіту рецептакуліти, які беруть участь у рифоутворенні поряд із кораловими поліпами підкласів Tabulata, Heliolithoidea та гідроїдними поліпами підкласу Stromatoporoidea (колоніальні форми, які відрізнялись масивним вапняковим скелетом особливої будови, рис. 242).
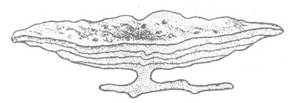
Рис. 242. Зовнішній вигляд колонії Stromatoporoidea (клас Hydrozoa) з ордовика
У прісних водоймах з’являються перші Oligochaeta. З членистоногих починається розквіт Trilobitomoipha; мечохвости представлені особливим рядом Chasmataspidida. У прісних водоймах з’являються велетенські (до 2—3 м завдовжки) хижі хеліцерові з ряду Eurypterida. Ракоподібні представлені тими самими групами, що й у кембрії. Серед молюсків з’являються ряд Mesogastropoda з черевоногих, клас Scaphopoda, кілька рядів двостулкових. Дуже швидкими темпами йде еволюція головоногих: процвітають Nautiloidea, з’являються Orthoceratoidea, велетенські Endoceratoidea та Actinoceratoidea. У всіх них черепашки були прямими.
Brachiopoda в цей період були однією з домінуючих груп. У ранньому ордовику в морях та прісних водоймах з’являються та дуже поширюються протягом періоду моховатки (Bryozoa). Спостерігається справжній розквіт голкошкірих: відомо близько 20 класів, у тому числі всі сучасні. Відомо багато видів крилозябрових (граптоліти), серед яких були бентосні, псевдопланктонні та планктонні форми.
Наприкінці періоду внаслідок геологічних і кліматичних змін вимирають деякі групи трилобітів, головоногих (Endoceratoidea) та голкошкірих.
Силурійський період характеризується появою перших наземних прибережних екосистем. Знижується різноманітність та чисельність рецептакулітів. З’являються вапнякові губки. Крім табулят, строматопорат та геліолітоідей, помітну роль у рифоутворенні починають грати ругози. З’являються морські пера (Pennatulacea). Мечохвости представлені сучасним рядом Limulida. Чисельність та різноманітність Trilobitomorpha знижується. У зв’язку з виходом на суходіл перших наземних рослин — рініофітів — виникають наземні екосистеми, в яких із безхребетних мешкають олігохети та скорпіони. У морях із молюсків з’являються Tentaculita, продовжують існувати майже всі ті самі підкласи головоногих, що й в ордовиці; з’являються види зі спіральною черепашкою. Силур — час розквіту морських лілей. Наприкінці періоду зникає більшість граптолітів.
Це теплий період, що характеризувався переважанням моря над суходолом. У північній півкулі кілька окремих платформ з’єднались в єдиний материк Лавразія.
Девонський період характеризується подальшим освоєнням живими організмами суходолу, в тому числі й безхребетними. Значні зміни у фауні спостерігаються і в морях. У цей період досягли значного розвитку деякі групи форамініфер з аглютинованими та вапняковими черепашками. З’являються реброплави. Рифоутворювачами були строматопорати, табуляти та ругози; вимирають геліоліїди. Видова різноманітність трилобітів значно знижується, з’являються морські павуки — Pantopoda. Серед зябродишних з’являються два ряди Branchiopoda: Conchostraca та відомий лише з девону Lipostraca, представлений одним родом — Lepidocaris.
Із цього періоду відомі кліщі та особливий ряд панцирних павуків (Soluta, рис. 243), тіло яких, на відміну від сучасних, було вкрите товстим хітиновим спинним щитом. На суходолі з’явллються перші трахейнодишні — ряд Collembola з класу Entognatha. У девоні відбувається бурхливий розвиток головоногих зі спіральною черепашкою (Nautiloidea). З’являється підклас Bactritoidea з прямою черепашкою, від якого виникають Ammonoidea та перші Belemnitida. Наприкінці періоду вимерли тентакуліти та багато класів голкошкірих (карпоідеї, морські пухирі тощо); продовжується розквіт морських лілей.
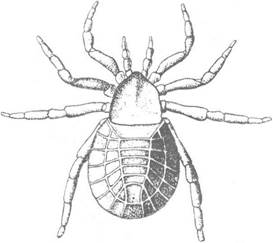
Рис. 243. Панцирний павук Crypto martus hindi з карбону
Протягом девону горотворчі процеси змінювались наступом моря, однак загалом наприкінці періоду існували два великих материки: північний — Атлантія та південний — Гондвана, що зумовило різку зональність клімату.
Кам’яновугільний період (карбон) відомий насамперед як час виникнення справжніх комах (Ectognatha), що пов’язано з освоєнням живими організмами більшої частини суходолу. Перші комахи відомі з середнього карбону. Це, з одного боку, первиннобезкрилі (рад Machilida), з іншого —кілька рядів крилатих комах. Найпримітивнішим із них був рад Protoptera. Це були комахи з гризучими ротовими органами, двома парами крил із сітчастим жилкуванням; розвиток, ймовірно, відбувався по типу архіметаболії. Вони мешкали на деревах та живились генеративними органами (спорангіями спорових чи стробілами голонасінних). Вважають, що від протоптер походять усі інші крилаті комахи; саме цей рад вимер у другій половині карбону. Види ряду Caloneurida були більш спеціалізовані порівняно з попереднім рядом: мали видовжене тіло й довгі ноги. Вони також живились генеративними органами деревних рослин, однак відкладали великі за розмірами яйця. Це свідчить про ембріонізацію розвитку, який у калоневрид, ймовірно, мав характер ге міметаболії. Види ряду Dictyoneurida мали колючосисний ротовий апарат та досить великі розміри (до 40 см у розмаху крил). Вони висмоктували вміст спорангіїв; це були предки клопів, рівнокрилих та деяких інших сучасних груп. У середньому карбоні виникли й такі хижаки, як бабки (Odonata). Деякі з них досягали 70 см у розмаху крил. Цікаво, що личинки бабок карбону вели наземний спосіб життя. У пізньому карбоні кількість радів комах значно збільшується. Це пояснюється тим, що комахи почали живитись рослинним опадом. З другої половини періоду відомо багато мешканців підстилки та поверхні ґрунту — детритофагів, що належали до рядів тарганових (Blattoidea), грилоблатид (Grylloblattida) (ці комахи живуть і в наш час; нагадують тарганів, однак передні крила не перетворені на надкрила) та деяких викопних груп. З кінця періоду відомі прямокрилі (Orthoptera), представлені на той час особливими групами хижаків. Усього на кінець карбону відомо 15 рядів комах. Цікаво, що у викопних комахах знайдено залишки паразитичних плоских червів, ймовірно, трематод. З цього часу відомі й ківсяки (Diplopoda).
У ґрунті та підстилці були поширені сапробіонті кліщі підряду Acariformes; більш спеціалізовані форми (фітофаги, хижаки, паразити тощо) появились значно пізніше. У карбоні виникають деякі групи хижих павукоподібних: справжні павуки (Aranei) (панцирні павуки вимирають наприкінці періоду), косарики (Opiliones), несправжні скорпіони (Pseudoscorpiones), сольпуги (Solifugae) та особливий карбоновий ряд Phalangiotarbi.
У цей час суходіл освоїли і черевоногі молюски — виникає підклас легеневих (Pulmonata).
Досить великі зміни відбуваються і в населенні морів. Серед форамініфер з’являється і досягає розквіту підряд Fusulinida з вапняковими багатокамерними веретеноподібними черепашками складної будови до 20 мм завдовжки; їх залишки утворили так звані фузулінові вапняки. Серед губок домінують представники ряду Tetraxonida; знайдено спікули прісноводних форм із ряду Comacuspongida, що нагадують сучасних. На початку періоду вимирають останні Receptaculita. З коралів домінують Tabulata та Rugosa, а чисельність Stromatoporata різко падає. Знайдено залишки Nemathelmin- thes та Chaetognatha.
З трилобітів зберігається лише один ряд Rychopariida. Серед зябродишних з’являються клас Cephalocarida, ряди Anostraca та Mysidacea. Представники цих рядів перші серед ракоподібних перейшли до планктонного способу існування.
Наприкінці періоду з’являються Opisthobranchia; двостулкові молюски за чисельністю та видовою різноманітністю значно поступаються Brachiopoda. Виникають ксеноконхії. Серед головоногих домінують форми зі спіральною черепашкою з підкласу Nautiloidea; також досить поширені Bactritoidea і Ammonoidea; вимирають Orthoceratoidea, Actinoceratoidea та деякі інші.
Різноманітність беззамкових Brachiopoda зменшується, переважають замкові. Серед голкошкірих домінують Clinoidea, Blastoidea та архаїчні форми Echinoidea. Протягом періоду вимирає ряд класів голкошкірих.
Ранній та середній карбон відрізнявся м’яким кліматом та переважанням моря над суходолом; у пізньому карбоні море відступає, клімат стає суворішим; відзначено часткове зледеніння Гондвани. Лавразія та Гондвана зближуються.
Пермський період — час повного освоєння живими організмами суходолу та становлення сучасних меж біосфери. Основу рослинності становили вищі спорові та голонасінні. Це період бурхливої адаптивної радіації комах; личинки багатьох груп (одноденки, бабки, веснянки тощо) освоюють прісні водойми. У цей період виникають одноденки (Ephemeroptera) з перетворенням типу архіметаболії; багато рядів комах з неповним перетворенням (сіноїди — Psocoptera, рівнокрилі — Homoptera, трипси — Thysanoptera та ін.); виникають комахи з повним перетворенням (ряди твердокрилі — Coleoptera, сітчастокрилі — Neuroptera, волохо- крильці —Trichoptera тощо). Вимирають деякі ряди кам’яновугільного періоду (Caloneurida, Dictyoneurida та ін.). Наприкінці перму було відомо 26 рядів комах, з яких 20 існують і досі.
Серед комах-фітофагів у цей період переважали форми з колючо-сисними ротовими органами; досить численні групи з гризучим ротовим апаратом, які живились переважно генеративними органами рослин та пилком; листогризучі пермські комахи невідомі. Існувало багато видів, що живились опалим листям, а також гнилою деревиною (жуки). Серед пермських комах відомі й хижаки, що належали до різних рядів.
У морях продовжували домінувати фузулініди, які, однак, повністю вимерли наприкінці періоду. Основними рифоутворювачами були коралові поліпи Tabulata та Rugosa, які також вимерли в кінці періоду; у рифоутворенні брали участь і Bryozoa, Spongia тощо. Із Malacostraca з’являються ряди Euphausiacea та Decapoda.
Протягом періоду вимерли останні трилобіти та Euiypterida. У водоймах зростає різноманітність черевоногих із ряду Mesogastropoda (підклас Prosobranchia) та Opisthobranchia. З головоногих поширені Nautiloidea, Ammonoidea та Belemnitida. Вимирають хіоліти. Значно зменшується кількість видів моховаток внаслідок вимирання поширених у палеозої рядів — Cryptostomata, Cystoporida та Rhabdomezonida; також падає чисельність плечоногих, бо в кінці періоду вимирає ряд Ргоdutida, а видова різноманітність інших таксонів значно знижується.
У середині періоду дуже сильно зменшується різноманітність і чисельність морських лілей та вимирають Blastoidea; інші класи голкошкірих трапляються досить рідко.
Протягом пермського періоду існували два суперконтиненти — північний (Лавразія) та південний (Годвана), які наприкінці цього періоду утворили єдиний суперконіинент — Пангею. Це спричинило загальне похолодання та різку природну зональність; значна частина суходолу мала помірний або холодний клімат; натомість в екваторіальній області чітко виділялись субтропічний та тропічний пояси.
Мезозойська ера. Ця ера в еволюції безхребетних відзначається тим, що впродовж її сформувалась більшість сучасних рядів та значна частина родин і родів у різних групах.
Тріасовий період характеризується значними змінами як серед наземних, так і серед водяних безхребетних. У цей час з’являються такі відомі ряди комах, як напівтвердокрилі (Неmiptera), паличники (Phasmoptera), двокрилі (Diptera) та перетинчастокрилі (Hymenoptera). Вимирає кілька палеозойських рядів. Трофічні зв’язки комах стають різноманітнішими. З фітофагів переважають сисні форми (клопи та рівнокрилі), однак з’являються і листогризучі —паличники та деякі прямокрилі. Більшість комах із гризучим ротовим апаратом живилась, як і в Пермі, генеративними органами та пилком рослин (сітчастокрилі, перетинчастокрилі, жуки). Основними детритофагами були різноманітні таргани, в гнилій деревині розвивались личинки двокрилих і жуків. Хижаками були личинки сітчастокрилих, більшість прямокрилих, бабки і жуки. Прісні водойми освоїли личинки волохокрильців, бабок, одноденок, веснянок; з’являються водяні клопи та жуки.
У морях мешкали кілька рядів форамініфер, які в цей період не досягали високої чисельності. У цей час з’являється ряд Leptolida (клас Hydrozoa); винятково важливою є поява мадрепорових коралів (ряд Scleractinia), які, однак, були ще нечисленні. Вимирають конуляти — особливий підклас сцифоїдних. Основними рифоутворювачами були строматопорати та губки. З’являються нові групи ракоподібних — Соpepoda, Isopoda та Tanaidacea; Decapoda досягають досить великої різноманітності. Вимирає ряд Archaeostraca, відомий починаючи з кембрію. Чисельність і видова різноманітність Brachiopoda різко зменшується з кінця тріасу, і до нашого часу вони займають другорядне місце в морських екосистемах та мешкають на значних глибинах. Натомість двостулкові молюски освоюють мілководдя морів та прісні водойми: починається їх розквіт, який триває й до нашого часу. Серед головоногих різко падає чисельність наутилоідей, вимирають бакрітоідеї, натомість спостерігається розквіт амоноідей та белемнітів.
У тріасовий період існував єдиний суперконтинент — Пангея, з численними внутрішніми прісно- та солоноводними басейнами; клімат тоді був дуже суворим; крім тропічних і субтропічних регіонів були зони помірного й холодного клімату.
В юрський період продовжується адаптивна радіація комах. З’являється ряд вуховерток (Dermaptera). Комахи відіграють усе більшу роль у деструкції рослинних решток. З’являються перші споживачі деревини тільки-но загиблих рослин: рогохвости з перетинчастокрилих та жуки-златки. Личинки двокрилих комах (так звані грибні комарики — Mycetophiloidea) заселили плодові тіла шляпкових грибів. З’являються паразитичні комахи — їздці та деякі інші перетинчастокрилі; їх хазяями були переважно інші види комах. Серед фітофагів поширені не тільки сисні, але й листогризучі, представлені пильщиками з перетинчастокрилих. Хижі види відомі серед прямокрилих (домінуюча форма хижаків), личинок сітчастокрилих, скорпіонових мух, деяких двокрилих та жуків; високої різноманітності досягають бабки, представлені двома сучасними та одним вимерлим наприкінці періоду підрядами. У прісних водоймах мешкають личинки багатьох рядів комах (одноденки, бабки, двокрилі, волохокрильці та ін.), клопи та жуки; усі вони були хижаками, що живились переважно різноманітними ракоподібними, дуже поширеними в прісних водоймах (переважно Conchostraca та Copepoda).
З юрського періоду відомі перші залишки Zoomastigophoгеа, Phytomastigophorea та Ciliophora. У морях знайдено медуз із сучасних рядів Semeostomea та Cubomedusae. Головними рифоутворювачами періоду були вапнякові губки, стромато- порати, моховатки і мадрепорові корали, для яких почався період розквіту.
У морях достатньо поширені як бентосні, так і планктонні ракоподібні. Спостерігалась значна видова різноманітність двостулкових та черевоногих молюсків. Дуже численні амоноідеї (бентосні й плаваючі форми); ці хижаки мешкали в прибережних водах на глибинах, що не перевищували 200 м. З наутилоідей залишається лише один ряд —Nautilida, представлений багатьма видами. Белемніти — плаваючі хижаки океанських просторів — досягають найбільшої різноманітності; у цей період від них виникли кальмари та каракатиці. Амоноідеї — також одна з домінуючих груп.
Продовжується скорочення числа видів Brachiopoda, вимирає багато груп цих тварин, і на кінець періоду об’єм типу нагадує сучасний, залишаються три ряди: Lingulida, Craniida, Terebratuhda. Серед голкошкірих домінують правильні морські їжаки, однак відомі й неправильні; виникає три ряди цих тварин, які існують і зараз.
Клімат юри був тепліший, ніж клімат тріасу; це був час розпаду Пангеї спочатку знову на Лавразію та Гондвану, а потім — на дрібніші континенти (наприклад, відділення від Гондвани Австралії). Були поширені континентальні мілководні теплі моря.
Крейдяний період характеризується насамперед біоцено- тичною кризою, яка відбулась у середині крейди та охопила наземні й прісноводні екосистеми. Її причиною була зміна екосистем, основою яких були мезозойські голонасінні (мезофіт), угрупуваннями, створеними покритонасінними та деякими групами хвойних (кайнофіт). Це призвело до вимирання під час кризи значної кількості таксонів тварин. Це вимирання, пов’язане зі зникненням місць існування, не було компенсоване виникненням нових таксонів і знизило кількість родин комах приблизно на 10 %. Під час формування нових екосистем з покритонасінними йшли процеси паралельної еволюції комах і квіткових рослин, що призвело до значного зростання кількості таксонів комах. Вимерли передусім групи фітофагів, що живились мезозойськими голонасінними, їх паразити та хижаки. Натомість у пізній крейді починається розвиток комах-запилювачів: деяких груп жуків, бджолиних з перетинчастокрилих, вищих мух; у цей час з’являються метелики (Lepidoptera), личинки яких живляться переважно листям та іншими органами квіткових і хвойних; виникають суспільні комахи, найважливішими з яких були терміти (Isoptera) та мурашки (Formicoidea); вони відігравали дуже важливу роль у ґрунтоутворенні, а терміти, крім того, — ще й в початкових стадіях деструкції мертвої деревини. З’являються богомоли — спеціалізовані хижаки, кровососи та паразити хребетних — мокреці з двокрилих (Ceratopogonidae), воші (Anopium) та блохи (Aphaniptera). Комплекси водяних комах ранньої крейди подібні до юрських; у пізній крейді спостерігається різке збіднення фауни водяних комах внаслідок змін характеру самих водойм; поряд із хижаками з’являються й фільтратори (личинки багатьох двокрилих тощо). Слід сказати, що в другій половині крейди закінчилось формування всіх рядів комах, які представлені й у сучасній фауні. З цього періоду відомі також перші Tardigrada.
У морях з’являються нові групи форамініфер (Heterohelicida, Nummulitida), які поряд з більш давніми таксонами (RotaUida, Lagonida, BuUmulida) досягають дуже високої чисельності та видової різноманітності; залишки їх черепашок утворюють поклади крейди, від якої й пішла назва періоду. Усі ці таксони існують і в наш час. Рифоутворювачами були ті самі групи, що і в юрський період.
Серед черевоногих молюсків виникає ряд Neogastropoda (передньозяброві); продовжується утворення нових таксонів Pulmonata. Серед двостулкових досягає великої різноманітності ряд Rudista, який виник ще в пізньому юрі; вони мешкали на мілководдях континентальних морів і вимерли наприкінці крейди. Інші групи Bivalvia досягли значної чисельності. Амоноідеї досягають максимальної видової різноманітності в ранній крейді, а в пізній число їх таксонів зменшується, і вони повністю вимирають у кінці періоду. Домінують белемніти, які, однак, майже повністю зникають наприкінці крейди; досить поширені каракатиці та кальмари; виникають восьминоги. У кінці періоду вимирає понад 50 % рядів і родин морських їжаків.
Протягом періоду Лавразія та Гондвана розпадаються на окремі континенти; загалом площа суходолу була менша, ніж зараз, а клімат значно тепліший.
Кайнозойська ера. На початку цієї ери серед безхребетних існувало ще багато мезозойських груп; протягом її фауна стає дедалі ближчою до сучасної.
Палеогеновий період характеризується становленням сучасних груп комах на рівні підрядів та частково родин. Досягають розквіту всі групи запилювачів та сисні й гризучі фітофаги, які живляться будь-якими органами рослин, їх хижаки та паразити. Виникають різноманітні некро- та копрофаги, що живляться трупами і фекаліями ссавців та птахів (коротковусі двокрилі, жуки-гробарики, гнойовики тощо). Комплекс кровососів збагачується мошками, москітами, кровосисними комарами, гедзями; з’являються оводи. Прісноводні водойми освоюють квіткові рослини, що збільшує біопродуктивність цих екосистем; тому багато груп комах знову заселює ці водойми, і їх фауна стає подібною до сучасної.
У палеогені з’являються кровосисні іксодові кліщі та гамазові кліщі —переважно мешканці ґрунтів. З найпростіших з’являються черепашкові амеби (Testacealobosia). У морях із форамініфер найпоширеніші нумуліти; черепашки деяких з них досягали 16 см у діаметрі; поряд з ними високої чисельності досягають і інші групи, відомі з часів крейди; наприкінці періоду видова різноманітність форамініфер значно зменшується.
Вимирають строматопорати; основними рифоутворювачами, як і в наш час, були мадрепорові корали.
Серед ракоподібних виникають Cladocera, які відразу ж стають суттєвим компонентом планктону як прісних, так і морських водойм; з’являються також бокоплави (Amphipoda). Продовжується розквіт черевоногих та двостулкових. Белемніти представлені лише однією родиною, яка вимирає наприкінці періоду; поширені також кальмари та каракатиці, трапляються й восьминоги. Починається новий розквіт морських їжаків.
Протягом періоду горотворчі процеси змінювались наступом морів, що призводило до змін клімату, який, однак, був тепліший ніж зараз. Існувало кілька континентів, наприклад, Атлантія (з’єднання Європи з Північною Америкою), Індія, Ангарія, Африка (без півночі, де було море), Південна Америка тощо.
Неогеновий період. Ще в кінці палеогену почали виникати трав’янисті біоценози (степи тощо), які остаточно сформувалися в неогені. Ці екосистеми заселяли специфічні комплекси комах (хортобіонти). По-перше, це травоїдні групи, чільне місце серед яких зайняли саранові, та їх паразити і хижаки. По-друге, це копрофаги, некрофаги, кровососи і паразити степових ссавців та птахів. По-третє, в умовах різнотрав’я утворюються специфічні до певних рослин запилювачі, насамперед, бджолині.
У морях фауна безхребетних близька до сучасної: дуже знижується кількість нумулітів; зростає роль планктонних форамініфер, частина яких вимерла наприкінці періоду.
Протягом неогену обриси материків стають переважно подібними до сучасних; встановились кліматичні зони — від тундри до тропіків. Кожна з цих зон має свій комплекс специфічних видів, що привело до загального збільшення видової різноманітності наземних безхребетних. Протягом періоду спостерігалось кілька зледенінь, з яких саме значне було в Південній півкулі 11 млн років тому, коли більша частина Австралії, Нова Зеландія та Патагонія знаходились під шаром криги і були заселені наново після кінця зледеніння.
Антропогеновий період характеризується кількома зледеніннями та появою антропогенного фактора. Фауна безхребетних протягом періоду істотно не відрізняється від фауни кінця неогену. Господарча діяльність людини загрожує стійкості біосфери в цілому та веде до вимирання багатьох видів, у тому числі й безхребетних.