Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Коферменты - особые природные специализированные реагенты
Липоевая кислота и окислительное декарбоксилирование а-кетокислот
Ферментативная функция
Окислительное декарбоксилирование а-кетокислот включает расщепление кетокислота с образованием СО2 и присоединение остающейся ацильной группы к СоА:
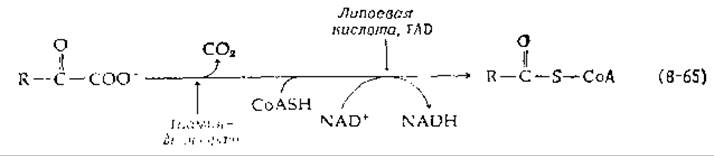
NAD+ выступает в этой реакции в роли окислителя. Реакция катализируется полиферментным комплексом с мол. весом ~1∙106—9∙106 в зависимости от биологического источника и от типа субстрата [137— 139]. Отдельные системы дегидрогеназ кетокислот известны для пировиноградной, а-кетоглутаровой кислот и для а-кетокислот с разветвленной боковой цепью, метаболически образующихся из лейцина, изолейцина и валина. Наиболее детально изучены пируватдегидрогеназа и а-кетоглутарат—дегидрогеназа Е. coli. В обоих случаях комплекс можно разделить на три компонента. Одним из них является декарбоксилаза (называемая также дегидрогеназой), у которой диссоциирующим кофактором является тиаминдифосфат. Вторым компонентом является флавопротеид дигидролипоилдегидрогеназа, а третьим — содержащий липоевую кислоту «сердцевинный» фермент (core enzyme).
По данным электронной микроскопии, этот фермент обнаруживает поразительную октаэдрическую симметрию, которая была подтверждена рентгеноструктурным анализом. Сердцевинный фермент дигидролипоил-трансацетилаза из пируватдегидрогеназы имеет мол. вес ~1,7∙106 и состоит из 24 идентичных, по-видимому, субъединиц с мол. весом, равным 70 000. Каждая субъединица содержит связанный липоильный остаток [138, 139]. Структура этого фермента схематически показана на рис. 8-17. С «сердцевиной» связаны примерно 12 димерных субъединиц декарбоксилазы-дегидрогеназы с мол. весом 192 000 и 6 молекул димерного флавопротеида с мол. весом 112 000. По-видимому, 12 димеров декарбоксилазы-дегидрогеназы симметрично распределены по двенадцати ребрам трансацетилазного куба, а 6-димеров флавопротеида — по шести плоскостям куба. Вероятно, активные центры всех субъединиц плотно пригнаны друг к другу в областях контакта субъединиц, в которых возможно протекание последовательности каталитических реакций, указанных на рис. 8-18.
Уникальная функция липоевой кислоты состоит в окислении связанного с тиамином «активного альдегида» таким образом, чтобы при распаде комплекса с тиамином ацильный остаток, возникающий при окислительном декарбоксилировании кетокислота, присоединялся к дигидролипоевой кислоте. Поскольку известно, что липоевая кислота присоединена к субъединицам «сердцевинного» фермента, ясно, что при помощи «ножки» длиной 1,5 нм липоевая кислота дотягивается до тиаминдифосфатного центра в одной из декарбоксилазных субъединиц. Получив ацильную группу, она снова поворачивается к тому центру «сердцевинного» фермента, с которым связан СоА. Ацильная группа переносится на СоА с образованием дигидролипоевой кислоты, которая затем совершает поворот к третьей субъединице, где имеются FAD и дисульфид, готовые окислить ее вновь в липоевую кислоту.
Восстановленный флавин-дисульфидный фермент затем окисляется за счет NAD+ (рис. 8-18).
Высказывалось предположение о наличии механизма, отличного от того, который показан на рис. 8-18. Феррицианид, который в системе пируватдегидрогеназы может замещать NAD+в качестве окислителя, также способен неферментативно окислять связанный тиамином активный альдегид в 2'-ацетилтиамин:
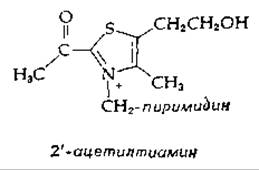
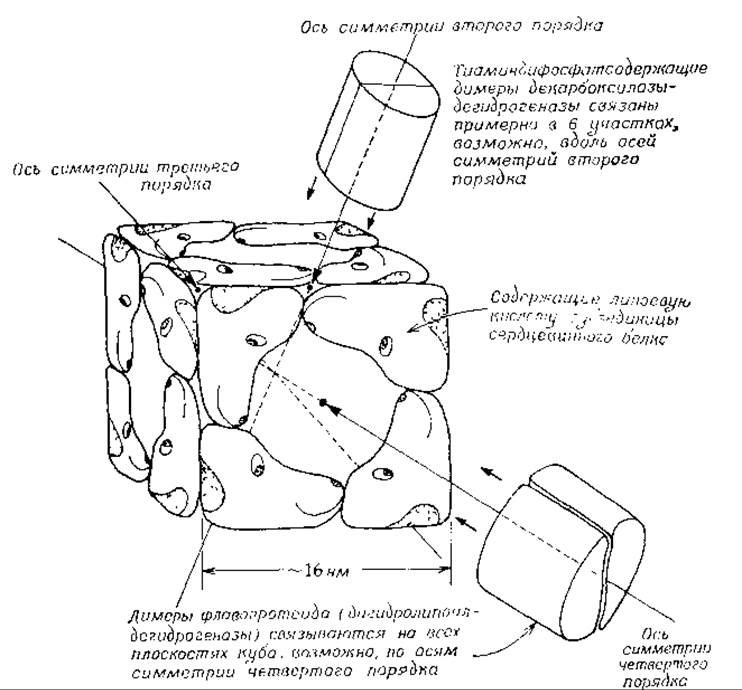
РИС. 8-17. Схема расположения 24 субъединиц в транссукцинилазном «ядре» а-кетоглутаратдегидрогеназы Е. coli. Для получения наблюдаемой кубической симметрии субъединицы «сердцевины» должны быть сгруппированы в тримеры по углам куба с образованием структуры, имеющей три поворотные оси симметрии четвертого порядка, четыре оси симметрии третьего порядка и шесть осей симметрии второго порядка (симметрия 4:3:2). Однако (см. текст) симметрия не совершенна: только в половине из всех субъединиц содержится связанная липоевая кислота и имеется только шесть декарбоксилазных димеров. В случае пируватдегидрогеназы Е. coli каждая трансацетилазная субъединица содержит липоевую кислоту и в комплекс входят 12 декарбоксилазных димеров.
Ацетильная группа этого соединения обладает высокой способностью к переносу. Таким образом, не исключена возможность, что липоевая кислота сперва окисляет активный альдегид до связанного с тиамином ацильного производного, а затем во второй стадии акцептирует ацильную группу по реакции нуклеофильного замещения. Однако этот механизм не позволяет объяснить уникальную роль липоевой кислоты в окислительном декарбоксилировании.

РИС. 8-18. Последовательность реакций, катализируемых дегидрогеназами а-кетокислот. Субстрат и продукт указаны в рамках, а путь превращения окисляемой кетокислоти отмечен жирными стрелками. «Головка» липоевой кислоты, способная к повороту, показана относительно точки присоединения к «сердцевиниой» субъединице.
Активность пируватдегидрогеназного комплекса тканей многих млекопитающих частично контролируется механизмом фосфорилирование — дефосфорилирование [138, 139а]. Фосфорилирование субъединицы декарбоксилазы (дегидрогеназы) ATP-зависимой киназой приводит к образованию неактивного фосфофермента. Особая фосфатаза реактивирует дегидрогеназу (рис. 6-15).
Наряду с зависимой от липоевой кислоты пируватдегидрогеназой, клетки Е. coli содержат пируватоксидазу — растворимый флавопротеид, который действует совместно с мембраносвязанной системой переноса электронов, превращая пируват в ацетат и СО2. Механизм этого процесса пока неясен [1386].