БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 37. ВОЗБУДИМЫЕ МЕМБРАНЫ И СЕНСОРНЫЕ СИСТЕМЫ
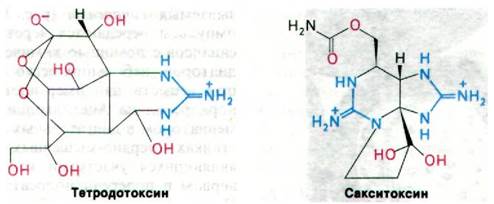
37.2. Тетродотоксин и сакситоксин блокируют натриевые каналы в мембранах аксонов нервных клеток
Тетродотоксин - сильнодействующий яд рыбы иглобрюха - блокирует проведение нервных импульсов вдоль аксона и в возбудимых мембранах нервных волокон, что вызывает паралич дыхания. Летальная доза для мыши составляет около 0,01 мкг. В связи с высокой специфичностью действия тетродотоксин с успехом используется при экспериментальных исследованиях. Он очень прочно (К ≈ 10-9 М) связывается с Na+-каналом и блокирует поток ионов натрия, не влияя при этом на К+-канал. Таким же действием обладает и сакситоксин, вырабатываемый одним из морских дино-флагеллят. Моллюски, питающиеся дино- флагеллятами, в особенности съедобные двустворчатые моллюски и мидии, тоже становятся ядовитыми. Так, одна мелкая мидия может содержать сакситоксин в дозе, достаточной чтобы убить 50 человек! Общая структурная особенность тетродотоксина и сакситоксина - наличие гуанидиновой группы (рис. 37.4). Эта положительно заряженная группа токсина взаимодействует с отрицательно заряженным карбоксилат- ионом в устье канала на внеклеточной стороне мембраны. В сущности, эти токсины являются конкурентными ингибиторами Na+.
Рис. 37.4. Блокаторы Na+-канала
Иглобрюх, считающийся в Японии деликатесом
Тетродотоксин и сакситоксины в силу своей специфичности и высокого сродства к Na+-каналу оказались ценнейшими средствами анализа. Так, измерением связыва-
ния меченого тетродотоксина с высокой удельной радиоактивностью определяли плотность Nа+-каналов в различных возбудимых мембранах. Немиелинизированные нервные волокна, которые лишены изоляционного слоя миелина, обычно характеризуются низкой плотностью Nа+-каналов - порядка 20 на 1 мкм2. В мембранах таких аксонов Na+-каналы отделены друг от друга расстоянием 2000 А. Что касается миелинизированных нервных волокон, то в специфических участках, называемых перехватами Ранвье, плотность Na+-каналов, напротив, достигает очень высоких значений - порядка 10 на 1 мкм2. Перехваты Ранвье, расположенные на аксоне с интервалом 2 мм, это единственные участки, в которых мембрана аксона миелинизированного нерва соприкасается с внеклеточной жидкостью. Участки мембраны между перехватами Ранвье содержат очень мало каналов и не участвуют в проведении. Потенциал действия перескакивает от перехвата к перехвату, вследствие чего импульс проводится быстрее и эффективнее, чем в немиелинизированном волокне. Наличие 104 каналов на 1 мкм2 в перехвате Ранвье означает, что значительная часть поверхности мембраны в этой области занята Na+-каналами.
Специфичность связывания тетродотоксина с Na+-каналами была использована также при их выделении и очистке. Для этого интегральные мембранные белки возбудимых мембран солюбилизировали с помощью детергента, после чего разделяли на ионообмeннике. Связывание тетродотоксина с солюбилизированным Na+-каналом позволило количественно определять канал в процессе очистки. Выделенный таким путем Na+-канал оказался белком массой 230 кДа, состоящим из субъединиц различных типов. Сложность устройства Na+-канала частично обусловлена, видимо, тем, что он является не только высокоизбирательной порой в мембране, но и содержит структуру, воспринимающую напряжение (сенсор напряжения). Заряженные группы этой структуры реагируют на изменение мембранного потенциала во время потенциала действия и передают информацию на ту часть канала, которая составляет пору в мембране. Действительно, до начала потока натрия через мембрану выявляется поток через канал (так называемый воротный ток), обусловленный движением заряженных групп в белке.
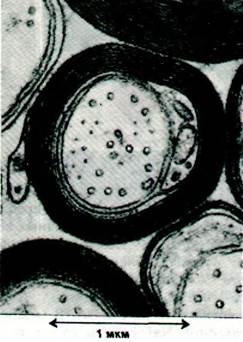
Рис. 37.5. Электронная микрофотография миелинизированного аксона из спинного мозга. Миелиновая оболочка аксона («обертка», образованная многочисленными слоями мембран) выполняет роль изолятора. Скорость проведения в миелинизированных нервах намного выше, чем в лишенных миелиновой оболочки нервах того же диаметра
37.3. Ацетилхолин является нейромедиатором
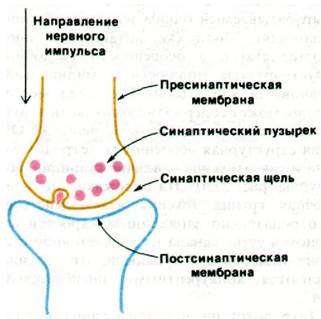
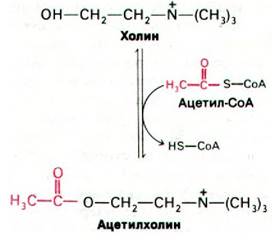
Взаимодействие между нервными клетками осуществляется в местах их соединения, называемых синапсами (рис. 37.6). Нервные импульсы передаются через большинство синапсов с помощью химических нейромедиаторов - небольших легко диффундирующих веществ, например, ацетилхолина или норадреналина. Ацетилхолин служит также медиатором в двигательных концевых пластинках (нервно-мышечных соединениях), являющихся участками контактов между нервом и поперечнополосатой мышцей. Пресинаптическая мембрана холинергического синапса (т. е. синапса, в котором в качестве нейромедиатора используется ацетилхолин) отделена от постсинаптической мембраны так называемой синаптической щелью шириной около 500 А. Окончание пресинаптического аксона наполнено синаптическими пузырьками (везикулами), содержащими ацетилхолин. Пришедший к синапсу нервный импульс вызывает высвобождение ацетилхолина в синаптическую щель. Далее молекулы ацетилхолина диффундируют через щель и достигают постсинаптической мембраны, где связываются со специфическими рецепторными молекулами. Это приводит к деполяризации постсинаптической мембраны, и далее волна деполяризации распространяется вдоль электрически возбудимой мембраны второй нервной клетки. Ацетилхолин гидролизуется ацетилхолинэстеразой, и постсинаптическая мембрана поляризуется вновь. Ацетилхолин синтезируется вблизи пресинаптического окончания аксона путем переноса ацетильной группы от ацетил-СоА на холин. Фермент, катализирующий эту реакцию, холин-ацетилтрансфераза (холин- ацетилаза). Далее часть образовавшегося ацетилхолина попадает в синаптические пузырьки, а часть остается в цитозоле. В одном холинергическом синаптическом пузырьке (обычно 400 А диаметром) содержится около 10000 молекул ацетилхолина.
Рис. 37.6. Схематическое изображение холинергического синапса
Изучению синаптической функции значительно способствовало выделение синаптосом из гомогената нервной ткани. Синаптосомы представляют собой пресинаптические окончания, образующие в процессе выделения замкнутые структуры. Они представляют собой мешочки из пресинаптической мембраны, содержащей митохондрии, цитозоль и синаптические пузырьки.
37.4. Ацетилхолин открывает в постсинаптической мембране каналы для катионов
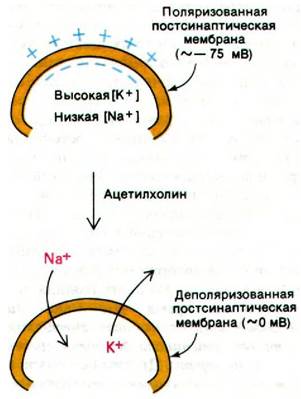
Потенциал покоя постсинаптической мембраны или мембраны двигательной концевой пластинки составляет примерно — —75 мВ. При взаимодействии ацетилхолина со специфическими рецепторами происходит резкое изменение проницаемости этих мембран (рис. 37.7). В течение 0,1 мс значительно возрастает проводимость как Nа+, так и К+, и возникает сильный ток Nа+ внутрь клетки и более слабый ток К+ из клетки. Ток Na+ в клетку приводит к деполяризации постсинаптической мембраны и инициирует возникновение потенциала действия в соседнем (постсинаптическом) нейроне или мышечном волокне. Ацетилхолин раскрывает катионные каналы только одного типа, характеризующиеся почти одинаковой проницаемостью для Nа+ и К+. Если поток Na+ при действии ацетилхолина оказывается большим, чем поток К+, то это обусловлено лишь большей крутизной электрохимического градиента концентрации Na+ по сравнению с К+.
Рис. 37.7. Ацетилхолин деполяризует постсинаптическую мембрану путем увеличения Na+ и К+ проводимости
Две молекулы ацетилхолина связываются с молекулой рецептора, вызывая при этом такие изменения конформации, которые открывают канал. Схематически кинетику процесса, согласующуюся с экспериментальными данными, можно описать уравнением
2А + R5 ⇄ A2R ⇄ A2R*,
где А - молекула ацетилхолина, R - закрытый канал, R* - открытый канал. За полупериод жизни открытого канала, равный всего лишь 1 мс, по нему проходит примерно 104 ионов. Продолжительное воздействие ацетилхолина на рецептор приводит к его десенсибилизации: канал закрывается, и реакция на ацетилхолин исчезает на длительный промежуток времени.