БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 26. ГЕНЕТИЧЕСКИЙ КОД Т ЗАВИСИМОСТЬ МЕЖДУ ГЕНАМИ И БЕЛКАМИ
26.2. Аминокислоты кодируются группами из трех оснований, начиная со строго определенной точки
Генетический код связывает последовательность оснований в ДНК (или в соответствующих транскриптах) и последовательность аминокислот в белках. К 1961 г. благодаря экспериментам Фрэнсиса Крика, Сиднея Бреннера (Francis Crick, Sydney Brenner) и других исследователей были установлены следующие свойства генетического кода.
1. Чему равно кодирующее отношение? Поскольку в ДНК имеется четыре вида оснований, то при кодировании одной аминокислоты одним основанием могло бы кодироваться всего лишь четыре аминокислоты. При кодировании одной аминокислоты двумя основаниями кодировалось бы 16 аминокислот (4 • 4=16), а при кодировании тремя основаниями - 64 аминокислоты (4 • 4 • 4 = 64). Белки состоят из двадцати аминокислот основного набора. Из этого несложного подсчета было очевидно, что для кодирования одной аминокислоты, видимо, необходимы три или более оснований. Генетические эксперименты показали, что на самом деле одну аминокислоту кодирует группа из трех оснований. Эта группа оснований называется кодоном.
2. Является ли код перекрывающимся? В случае непрерывающегося триплетного кода каждая группа из трех оснований ко-
дирует только одну аминокислоту, тогда как в случае полностью перекрывающегося триплетного кода АБВ кодирует первую аминокислоту, БВГ-вторую, ВГД-третью и т.д.
Эту дилемму удалось решить путем определения последовательности аминокислот в мутантах. Предположим, что основание В мутировало в В'. Если код не перекрывается, изменится только одна аминокислота. При полностью перекрывающемся коде мутация В в В' приведет к изменению аминокислот 1, 2 и 3. Изучение последовательности аминокислот белка оболочки мутантов вируса табачной мозаики показало, что у этих мутантов измененной обычно оказывалась только одна аминокислота. Напомним также, что как уже говорилось при обсуждении аномальных гемоглобинов в гл. 5, и в этом случае у большинства мутантов происходило изменение только одной аминокислоты. Отсюда был сделан вывод, что генетический код не перекрывается:
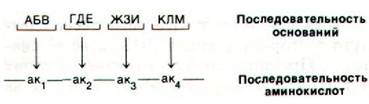
3. Как происходит правильное считывание группы из трех оснований? A priori одна из возможностей состоит в том, что одно из четырех оснований (оно обозначено Q) служит «запятой» между группами по три основания:
. . . QAБBQГДEQЖЗИQКЛMQ. . .
Оказалось, что это не так. Последовательность оснований читается последовательно, начиная со строго определенной точки:
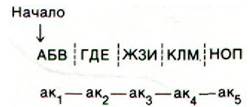
Запятых в коде нет. Предположим, что в результате мутации произошла делеция основания Ж:
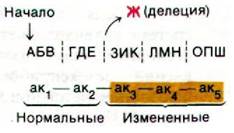
Первые две аминокислоты в этой полипептидной цепи будут нормальными, но остальная последовательность оснований будет прочитана неправильно, так как в результате делеции Ж произошел сдвиг рамки считывания. Теперь предположим, что между Е и Ж добавилось основание Ш:
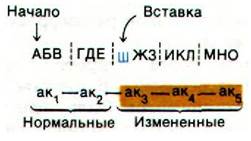
Эта вставочная мутация также нарушает рамку считывания, начиная с кодона аминокислоты 3. В действительности генетические исследования вставочных и делеционных мутантов позволили выяснить многие свойства генетического кода.
4. Как уже было сказано выше, существует 64 возможных триплета оснований и 20 аминокислот. Соответствует ли каждой из 20 аминокислот только один триплет, или некоторые аминокислоты кодируются более чем одним триплетом? Генетические исследования показали, что большинство из 64 триплетов кодируют аминокислоты. В последующих биохимических исследованиях было установлено, что 61 триплет из 64 кодирует определенные аминокислоты. Таким образом, для большинства аминокислот имеется более одного кодового слова. Другими словами, генетический код вырожден.
26.3. Расшифровка генетического кода: синтетические РНК могут служить информационными РНК
Каково соотношение между 64 видами кодовых слов и 20 аминокислотами? В принципе на этот вопрос можно получить прямой ответ, если сравнить последовательность аминокислот в каком-либо белке с соответствующей последовательностью оснований его гена или мРНК. Однако в 1961 г. этот подход был совершенно недоступен, так как о последовательностях оснований в генах и молекулах мРНК ничего не было известно. Тогда казалось, что проблема генетического кода не может быть решена в близком будущем, но внезапно ситуация изменилась. Маршалл Ниренберг (Marshall Nirenberg) обнаружил, что добавление полиуридилата [poly(U)] в бесклеточную систему синтеза белка приводит к синтезу полифенилаланина. Очевидно, poly(U) выполнил функцию информационной РНК. Первое кодовое слово было расшифровано: UUU кодирует фенилаланин. Этот замечательный эксперимент указал путь к полной расшифровке генетического кода.
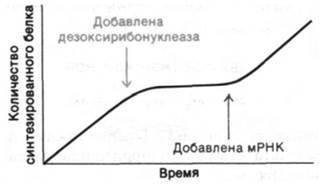
Обсудим более подробно этот эпохальный эксперимент. В качестве двух основных компонентов были использованы бесклеточная система, активно синтезировавшая белок, и синтетический полирибо- нуклеотид, сыгравший роль информационной РНК. Бесклеточная система синтеза белка была получена из Е. coli следующим образом. Бактериальные клетки осторожно разрушали, перемалывая с тонко измельченным порошком окиси алюминия, чтобы получить клеточный сок. Затем обрывки клеточной стенки и клеточной мембраны удаляли центрифугированием. В результате получали экстракт, содержащий ДНК, мРНК, тРНК, рибосомы, ферменты и другие клеточные компоненты. При добавлении ATP, GTP и аминокислот в этой бесклеточной системе синтезировался белок. По крайней мере одна из добавленных аминокислот была радиоактивной, что давало возможность обнаружить ее включение в белок. Эту смесь инкубировали при 37°С примерно в течение 1 ч. Затем добавляли трихлоруксусную кислоту, чтобы остановить реакцию и осадить белки. При этом свободные аминокислоты оставались в надосадочной фракции. Осадок промывали и просчитывали его радиоактивность, определяя таким образом сколько меченой аминокислоты включилось в новосинтезированный белок. Важная особенность этой системы состоит в том, что синтез белка можно остановить добавлением дезоксирибонуклеазы, разрушающей матрицы для синтеза новых мРНК. Поскольку та мРНК, которая уже имелась в смеси ко времени добавления дезоксирибонуклеазы, лабильна, синтез белка можно прекратить в течение нескольких минут. Затем Нирен- берг обнаружил, что синтез белка возобновляется при добавлении неочищенной фракции мРНК. Таким образом, в руках Ниренберга была бесклеточная система синтеза белка, зависящая от добавления мРНК.
Другим важнейшим компонентом в этом эксперименте был синтетический полири- бонуклеотид poly(U). Poly (U) синтезировали с помощью полинуклеотидфосфори- лазы - фермента, открытого в 1955 г. Марианной Грюнберг-Манаго и Северо Очоа (Marianne Grunberg-Manago, Severo Ochoa). Этот фермент катализирует синтез полири- бонуклеотидов из рибонуклеозиддифосфатов:
(РНК)n + Рибонуклеозиддифосфат ⇄ (РНК)n+1 + Рi
Полинуклеотид-фосфорилаза и РНК-полимераза катализируют совершенно различные реакции. В приведенной реакции активированными предшественниками служат рибонуклеозиддифосфаты, а не три- фосфаты. Продукт реакции - ортофосфат, а не пирофосфат. Следовательно, равновесие в реакции не может быть сдвинуто вправо в результате гидролиза пирофосфата. В самом деле, in vivo равновесие сдвинуто в сторону распада РНК, а не ее синтеза. Принципиальное отличие состоит в том, что полинуклеотид-фосфорилаза не использует матрицы. Состав РНК, синтезированной этим ферментом, определяется соотношением рибонуклеотидов в инкубационной смеси, а последовательность близка к случайной. Благодаря этому полинуклеотид-фосфорилаза оказалась ценнейшим инструментом в экспериментах по расшифровке генетического кода. Например, poly(U) синтезировали, инкубируя в присутствии фермента раствор высокой концентрации UDP. Сополимеры двух рибонуклеотидов, например, U и А, со случайной последовательностью готовили, также инкубируя UDP и АТР с этим ферментом.
Рис. 26.3. Синтез белка в бесклеточной системе останавливается через несколько минут после добавления дезоксирибонуклеазы и возобновляется при добавлении мРНК
Различные синтетические рибонуклеотиды вводили в бесклеточную систему синтеза белка и измеряли включение меченного 14С L-фенилаланина. Результаты оказались поразительными:

Тот же эксперимент был проделан с различными С-аминокислотами в каждой инкубационной смеси. Оказалось, что poly(А) вызывает синтез полилизина, a poly (С) - синтез полипролина. Так было расшифровано три кодовых слова:

Кодовое слово GGG невозможно было расшифровать таким же образом, потому что poly (G) не работает в качестве матрицы. Возможно, это объясняется тем, что он образует трехцепочечную спиральную структуру. Полирибонуклеотиды, образующие протяженные участки с упорядоченной структурой, не эффективны в качестве матриц для синтеза белка.