БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 28. РЕГУЛЯЦИЯ ВЫРАЖЕНИЯ ГЕНА В ФЕНОТИПЕ
28.2. Открытие регуляторного гена
Ключом к изучению механизма индукции β-галактозидазы стало открытие того факта, что под действием всех исследованных индукторов количество пермеазы и трансацетилазы возрастало прямо пропорционально количеству β-галактозидазы. Дальнейший прогресс был достигнут при изучении мутантов. Оно показало, что в-галакто- зидаза, пермеаза и трансацетилаза кодируются тремя генами - z, у и а соответственно - которые расположены друг за другом. Были получены мутанты, утратившие способность к синтезу одного из этих белков. Например, генотип z-y+a+ обозначает, что у данного мутанта нет β-галактозидазы, но имеются в нормальном количестве пермеаза и трансацетилаза. Наибольший интерес представляет класс мутантов, у которого мутацией затронуты все три белка. Такие конститутивные мутанты синтезируют без всякого индуктора большие количества в-галактозидазы, пермеазы и трансацети- лазы. Франсуа Жакоб и Жак Моно (Francois Jacob, Jacques Monod) пришли к выводу, что скорость синтеза этих трех белков зависит от какого-то общего элемента, отличного от генов, кодирующих их последовательности. Ген этого общего регуляторного элемента был обозначен ¡. Индуцибельные бактерии дикого типа имеют генотип i+z+ y+a+, а конститутивные мутанты по лактозным генам - генотип i-z+y+a+.
Каким образом осуществляется влияние гена; на скорость синтеза белков, кодируемых генами z, у и а? Проще всего было предположить, что ген; кодирует синтез некоего компонента цитоплазмы, названного репрессором, которого либо совсем нет в i-клетках, либо он там неактивен. Эта идея была проверена в ряде изящных генетических экспериментов с частично диплоидными бактериями, имевшими два набора генов лактозной области. Один набор содержался в бактериальной хромосоме, а второй - в половом факторе F', введенном в клетку при конъюгации. Например, был получен диплоид i+z+ Fi-z+. У этого диплоида гены i+ z- находятся в хромосоме, а гены i-z+ в эписоме. Является ли этот диплоид индуцибельным или конститутивным в отношении β-галактозидазы? Другими словами, может ли ген ¡+ бактериальной хромосомы подавлять экспрессию гена z+, расположенного в эписоме? Эксперимент дал совершенно четкий результат: диплоид индуцибелен, а не конститутивен. Такой же результат был получен и в отношении диплоида i-z+/Fi+z-. Следовательно, ген; кодирует способный к диффузии репрессор.
28.3. Оперон - единица координированной генетической экспрессии
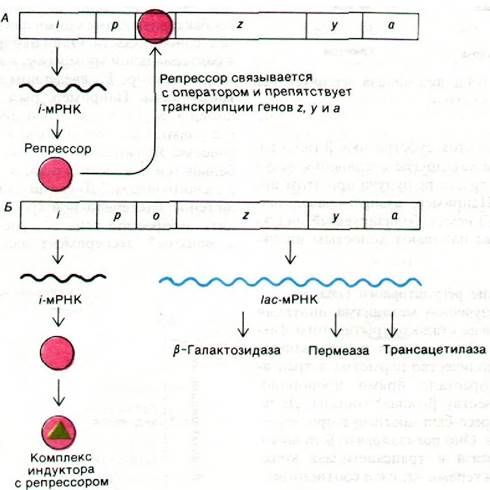
На основе только что описанных экспериментов Жакоб и Моно постулировали модель оперона, объясняющую регуляцию белкового синтеза. Генетические элементы этой модели - регуляторный ген, операторный ген и набор структурных генов (рис. 28.3). Регуляторный ген продуцирует репрессор, который может взаимодействовать с операторным геном. В дальнейшем выяснилось, что репрессор представляет собой белок. Операторный ген расположен рядом по соседству со структурными генами, которые он контролирует. Связывание репрессора с операторным геном препятствует транскрипции структурных генов. Операторный ген в совокупности со структурными генами, рядом с которыми он расположен, называется опероном. В случае лактозного оперона ген i - регуляторный ген, ген о - ген-оператор, а гены z, у и а - структурные гены. Кроме того, существует еще промоторный участок (обозначаемый символом р) для связывания РНК-полимеразы. Этот участок инициирования транскрипции расположен перед операторным геном. Индуктор, например, изопропилтиогалактозид (ИПТГ), связывается с репрессором и тем самым нарушает его взаимодействие с операторным геном. Теперь гены z, у и а могут транскрибироваться. При этом образуется одна длинная молекула РНК, кодирующая все три белка (рис. 28.4). Молекулу мРНК, кодирующую более одного белка, называют полицистронным (или полигенным) транскриптом.
Рис. 28.3. Карта лактозного оперона и его регуляторного гена. В карте масштаб не соблюден: участки р и о в действительности гораздо меньше, чем кодирующие участки генов
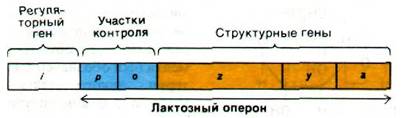
Рис. 28.4. Схема лактозного оперона в репрессированном (А) и индуцированном (Б) состояниях
28.4. lас-Репрессор - тетрамерный белок
При выделении репрессора лактозного опeрона (/ас-репрессор) была использована его способность связываться с ИПТГ. Уолтер Гилберт и Бенно Мюллер-Хилл (Walter Gilbert, Benno Muller-Hill) установили, что /ас-репрессор - белок, который связывается с ДНК, содержащей /ас-оперон, но не связывается ни с одной другой ДНК. Как и предполагалось, ИПТГ подавляет связывание /ас-репрессора с /ас-операторной ДНК. Клетка Е. co/i дикого типа содержит всего около десяти молекул /ас-репрессора. При очистке репрессора возникли трудности, связанные с тем, что он составляет всего 0,001% общего содержания белка. Однако существуют ieg-мутанты, имеющие, по-видимому, более эффективный промотор гена i. У этих мутантов образуется гораздо больше /ас-репрессора. Количество /ас-репрессора увеличивается еще сильнее, если использовать трансдуцирующие фаги, несущие область /ас. Зараженные таким фагом клетки Е. coli содержат около 20 000 молекул репрессора (примерно 2% всего белка). Это прекрасный исходный материал для выделения /ас-репрессора.
Рис. 28.5. Электронная микрофотография /ас-репрессора, связанного с ДНК, содержащей lас-оператор

Репрессор - тетрамер из идентичных субъединиц с массой 37 кДа, каждая из которых имеет один участок связывания с индуктором. Константа диссоциации ИПТГ составляет примерно 10-6 М. Репрессор очень прочно и быстро связывается с оператором. Константа диссоциации комплекса репрессор-оператор составляет примерно 10-13 М. Такое высокое сродство обусловлено тем, что в клетке Е. coli дикого типа содержится всего несколько молекул репрессора. Константа скорости ассоциации поразительно высока - 7 • 109 м-1 • с-1. Из этого следует, что репрессор находит операторный участок, диффундируя вдоль молекулы ДНК (одномерный поиск), а не ищет его, находясь в водной среде (трехмерный поиск)1.
1 Недавно проведенные исследования показали, что ситуация намного сложнее: поиск оператора репрессором включает оба указанных механизма и в очень большой степени зависит от ионного состава среды. Прим. перев.