БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 28. РЕГУЛЯЦИЯ ВЫРАЖЕНИЯ ГЕНА В ФЕНОТИПЕ
28.8. Транскрипция триптофанового оперона регулируется и аттенюатором, и оператором
Еще один регуляторный элемент был открыт Чарльзом Янофски (Charles Yanofsky) и его коллегами при изучении триптофано- вого оперона E. coli. Транскрибируемая с этого оперона мРНК длиной 7kb кодирует пять ферментов, превращающих хоризмат в триптофан (разд. 21.9). Эти пять белков синтезируются при трансляции полици- стронной trp-мРНК последовательно, координированно и в эквимолярных количествах. Трансляция начинается раньше, чем заканчивается транскрипция. trp-мРНК синтезируется примерно за 4 мин и затем быстро разрушается. Короткое время жизни trp-мРНК, составляющее всего около 3 мин, позволяет бактериям быстро реагировать на изменяющуюся потребность в триптофане. Е. coli может менять скорость образования ферментов биосинтеза триптофана более чем в 700 раз.
Как осуществляется эта регуляция? Первый уровень регуляции достигается путем взаимодействия специфического репрессора с trp-операторным участком ДНК. trp- репрессор - белок с массой 58 кДа, кодируемый геном trpR, удаленным от trp-оперо- на на довольно большое расстояние. Комплекс этого репрессора и триптофана прочно связывается с оператором, тогда как сам по себе репрессор с ним не связывается. Другими словами, триптофан является корепрессором. Мишень, на которую действует комплекс триптофана с репрессором, участок ДНК, обладающий симметрией второго порядка (рис. 28.11); и в этом случае симметрия играет важную роль во взаимодействии белка с ДНК. Этот операторный участок перекрывается с промоторным участком инициирования транскрипции. Таким образом, связывание trp-penpeccopa с оператором препятствует связыванию РНК-полимеразы с trp-промотором, и гены trp не транскрибируются.
Рис. 28.10. Схема trр-оперона. Показаны контрольные участки - промотор (р), оператор (о) и аттенюатор (а), а также гены, кодирующие лидерную последовательность (К), и ферменты пути биосинтеза триптофана (Е, Б, С, В и А)
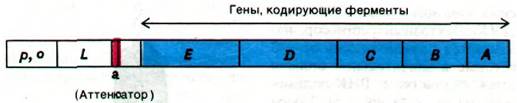
Рис. 28.11. Последовательность оснований trp-оператора. Ось симметрии второго порядка обозначена зеленым. Пара оснований, отмеченная +1,- начало транскрибируемой части оперона

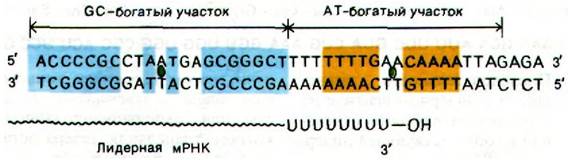
В течение некоторого времени считалось, что ингибирование конечным продуктом каталитической активности первого ферментативного комплекса в пути биосинтеза триптофана (разд. 21.11) и система ингибирования транскрипции с участием репрессора и оператора - основные регуляторные системы биосинтеза триптофана. Эта точка зрения была неожиданно опровергнута, когда было обнаружено, что у некоторых мутантов с делециями между оператором и геном первого фермента (trpE) в опероне происходит повышенное образование trp-мРНК. К тому же анализ 5'-концевой последовательности trp-мРНК показал, что там имеется лидерная последовательность длиной 162 нуклеотида, расположенная перед инициирующим кодоном trpE. Затем оказалось, что делеции, повышающие содержание trp-мРНК, картируются в этой лидерной области, примерно в 30-60 нуклеотидах от начала гена trpE. Следующее поразительное наблюдение состояло в том, что при высокой концентрации триптофана образовывался транскрипт, содержащий всего 130 нуклеотидов лидерной последовательности; при нехватке же триптофана синтезировалась trp-мРНК длиной 7000 нуклеотидов, включающая полную лидерную последовательность. Отсюда Янофски сделал вывод, что транскрипция trp-оперона должна регулироваться участком контролируемой терминации, называемым аттенюатором. Он локализован между оператором и геном первого фермента пути биосинтеза триптофана. Этот участок терминации, регулируемый физиологическими условиями, подобен участкам терминации в конце других оперонов (разд. 25.15); он содержит GC-богатую последовательность и следом за ней АТ-богатый участок. Каждый из этих участков аттенюатора обладает симметрией второго порядка (рис. 28.12). Кроме того, терминированный лидерный транскрипт заканчивается несколькими U подряд.
Рис. 28.12. Последовательность оснований аттенюаторного участка Связанные симметрией второго порядка пары оснований GC-богатой области показаны синим цветом; такие же пары AT-богатой области показаны желтым цветом
Аттенюаторный участок дополняет оператор в регуляции транскрипции trp-генов. При изобилии триптофана инициация транскрипции блокируется в результате связывания комплекса триптофан-репрессор с оператором. По мере снижения концентрации триптофана в клетке репрессия снижается и начинается транскрипция. Однако некоторые молекулы РНК-полимеразы покидают матрицу, дойдя до аттенюатора, тогда как другие продолжают синтезировать полную trp-матрицу. По мере исчерпания триптофана увеличивается доля молекул РНК- полимеразы, проходящих через аттенюаторный участок.
28.9. Аттенюация опосредуется трансляцией лидерной мРНК
Каким образом аттенюаторный участок trp- оперона улавливает концентрацию триптофана в клетке? В решении этого вопроса особенно важную роль сыграли данные о том, что часть лидерной мРНК транслируется. Весьма существенно, что в 14-членном лидерном полипептиде (рис. 28.13) имеются остатки триптофана в положениях 10 и 11. Когда триптофан находится в избытке, синтезируется полный лидерный пептид. Но если триптофана не хватает, рибосома задерживается на двух расположенных тандемом кодонах UGG, поскольку в этом случае оказывается недостаточным содержание триптофанил-тРНК. Застрявшая рибосома каким-то образом меняет структуру мРНК, так что РНК-полимераза транскрибирует оперон за пределами аттенюаторного участка. Ключевой аспект этого регуляторного механизма состоит в том, что трансляция и транскрипция тесно сопряжены между собой. Рибосомы, транслирующие лидерную trp-мРНК, следуют непосредственно за молекулой РНК-полимеразы, транскрибирующей ДНК-матрицу. Исследования, проведенные в последние годы, показали, что застрявшая рибосома изменяет вторичную структуру мРНК: конформация, при которой основания спариваются, благоприятствуя тем самым терминированию транскрипции, изменяется таким образом, что РНК-полимераза проскакивает аттенюатор (рис. 28.14). Мы начинаем понимать, что молекулы нуклеиновых кислот, как и белковые молекулы, могут принимать различные конформации и что изменения конформации регулируются и имеют далеко идущие физиологические последствия.
Рис. 28.13. Последовательность аминокислот в лидерном пептиде trp и последовательность оснований в соответствующей лидерной мРНК

Рис. 28.14. Схематическое изображение аттенюации trp-оперона E. coli. Когда триптофан имеется в избытке (А), лидерный участок (обозначен цифрой 1) trp-мРНК полностью транслируется. Участок 2 взаимодействует с рибосомой, что позволяет основаниям участков 5 и 4 спариваться. Эта спаренная область каким-то образом сигнализирует РНК-полимеразе о том, что следует закончить транскрипцию. Если же триптофана не хватает (Б), участки 3 и 4 не взаимодействуют, так как рибосома застревает на trp-кодонах участка 1. Участок 2. взаимодействует с участком 3. вместо того, чтобы входить в рибосому, и в результате участки 3 и 4 не могут спариваться. Вследствие этого транскрипция продолжается

28.10. Аттенюаторный участок гистидинового оперона содержит семь гистидиновых кодонов подряд
В настоящее время известны еще два оперона биосинтеза аминокислот у E. coli, содержащих аттенюаторные участки. Фенилала- ниновый оперон и гистидиновый оперон, подобно триптофановому оперону, содержат регулируемые участки терминации перед первым геном, кодирующим фермент. И в этих случаях лидерная область перед участком терминации транслируется. Удивительна последовательность аминокислот в лидерном пептиде фенилаланинового оперона: 7 из 15 остатков - фенилаланины (рис. 28.15). Еще поразительнее лидерный пептид гистидинового оперона: он содержит семь остатков гистидина подряд. Очевидно, что эти лидерные мРНК предназначены, чтобы улавливать концентрации фенилаланина и гистидина. Если соответствующих аминоацилированных тРНК не хватает, трансляция лидера останавливается. Как уже обсуждалось выше на примере trр-оперона, считается, что застрявшая рибосома таким образом изменяет конформацию мРНК, что у нее происходит спаривание оснований. Это дает возможность РНК-полимеразе проскакивать аттенюаторный участок1. Присутствие семи последовательно расположенных кодонов гистидина в лидерной мРНК гистидинового оперона существенно увеличивает чувствительность этого детектора. Действительно, падение концентрации гистидил-тРНК на 15% вызывает троекратное увеличение числа молекул мРНК, транскрибируемых с этого оперона.
1 Автор противоречит собственному утверждению, что застрявшая рибосома разворачивает мРНК (см. предыдущий разд.); рибосома действительно способствует именно разворачиванию мРНК. Прим. перев.
Рис. 28.15. Последовательность аминокислот в лидерном пептиде и последовательность оснований соответствующего участка мРНК фенилаланинового (А) и гистидинового (Б) оперонов

Рис. 28.16. Электронная микрофотография фагов![]()
