БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 28. РЕГУЛЯЦИЯ ВЫРАЖЕНИЯ ГЕНА В ФЕНОТИПЕ
28.11. Репрессоры и активаторы детерминируют развитие умеренных фагов
Обратимся теперь к роли репрессоров и активаторов транскрипции в регуляции жизненного цикла бактериофага лямбда (![]() ). Зрелая вирусная частица состоит из линейной двухспиральной молекулы ДНК (48 kb), упакованной в белковую оболочку. Существует два пути развития вируса: он может разрушить клетку-хозяина или он может стать ее компонентом (отсюда и название - умеренный). При литическом пути развития происходит полное выражение (экспрессия) фаговых генов, что приводит к лизису бактерии и образованию примерно 100 вирусных частиц потомства. В другом случае развитие фага X может пойти по пути лизогенизации клетки, когда его ДНК становится ковалентно связанной с ДНК клетки-хозяина в строго определенном месте (сайт-специфическая интеграция). Этот процесс рекомбинации, в котором участвует кольцевая молекула ДНК фага
). Зрелая вирусная частица состоит из линейной двухспиральной молекулы ДНК (48 kb), упакованной в белковую оболочку. Существует два пути развития вируса: он может разрушить клетку-хозяина или он может стать ее компонентом (отсюда и название - умеренный). При литическом пути развития происходит полное выражение (экспрессия) фаговых генов, что приводит к лизису бактерии и образованию примерно 100 вирусных частиц потомства. В другом случае развитие фага X может пойти по пути лизогенизации клетки, когда его ДНК становится ковалентно связанной с ДНК клетки-хозяина в строго определенном месте (сайт-специфическая интеграция). Этот процесс рекомбинации, в котором участвует кольцевая молекула ДНК фага ![]() , мы обсудим ниже (разд. 30.16). Когда ДНК фага интегрирует с ДНК клетки-хозяина, большинство фаговых функций выключается. Фаговая ДНК в таком состоянии называется профагом, а клетка-хозяин, содержащая профаг - лизогенной бактерией. Профаг реплицируется в лизогенной бактерии как часть клеточной хромосомы обычно на протяжении многих поколений. В лизогенном состоянии литические функции молчат, но не теряются (рис. 28.17). Многие агенты, нарушающие нормальную репликацию ДНК в клетке-хозяине, индуцируют профаг, и он переходит на путь литического развития.
, мы обсудим ниже (разд. 30.16). Когда ДНК фага интегрирует с ДНК клетки-хозяина, большинство фаговых функций выключается. Фаговая ДНК в таком состоянии называется профагом, а клетка-хозяин, содержащая профаг - лизогенной бактерией. Профаг реплицируется в лизогенной бактерии как часть клеточной хромосомы обычно на протяжении многих поколений. В лизогенном состоянии литические функции молчат, но не теряются (рис. 28.17). Многие агенты, нарушающие нормальную репликацию ДНК в клетке-хозяине, индуцируют профаг, и он переходит на путь литического развития.
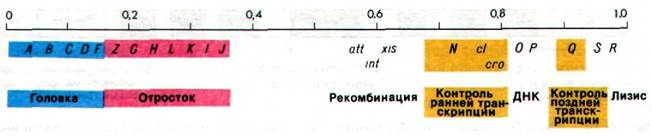
Рис. 28.17. Генетическая карта фага ![]() . Показаны лишь некоторые гены. После проникновения в бактериальную клетку линейная двухцепочечная ДНК переходит в кольцевую форму
. Показаны лишь некоторые гены. После проникновения в бактериальную клетку линейная двухцепочечная ДНК переходит в кольцевую форму
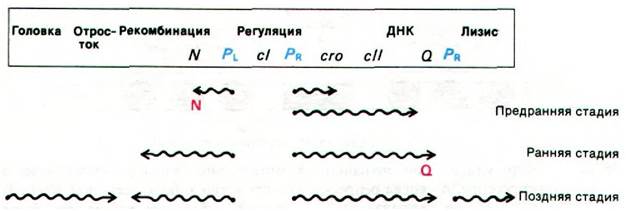
Вначале рассмотрим выражение генов фага ![]() при литическом пути. Цель развития - образование многочисленного потомства - достигается путем последовательной транскрипции вирусных генов. Вначале образуются белки, необходимые для репликации и рекомбинации ДНК, затем белки головки и отростка вирусной частицы и белки, необходимые для лизиса клетки-хозяина. Чрезвычайно важное значение имеет строгая очередность этих событий; преждевременное разрушение клетки-хозяина для вируса, конечно, невыгодно. Выражение генов при литическом развитии происходит в три стадии: предраннюю, раннюю и позднюю (рис. 28.18). На предранней стадии начинается синтез РНК с двух промоторов - PL и PR. Один из образующихся при этом транскриптов служит матрицей для синтеза белка N, которому принадлежит важнейшая регуляторная роль. В отсутствие белка N предранние транскрипты заканчиваются на одном из двух участков терминации. Белок N препятствует терминированию транскрипции в этих участках и обеспечивает таким образом дальнейшее выражение генов фага X. Белок N включает раннюю стадию. В это время синтезируются белки, необходимые для репликации ДНК фага и рекомбинации. Кроме того, на ранней стадии транскрибируется ген Q. Белок Q - еще один важный регуляторный элемент выражения генов фага
при литическом пути. Цель развития - образование многочисленного потомства - достигается путем последовательной транскрипции вирусных генов. Вначале образуются белки, необходимые для репликации и рекомбинации ДНК, затем белки головки и отростка вирусной частицы и белки, необходимые для лизиса клетки-хозяина. Чрезвычайно важное значение имеет строгая очередность этих событий; преждевременное разрушение клетки-хозяина для вируса, конечно, невыгодно. Выражение генов при литическом развитии происходит в три стадии: предраннюю, раннюю и позднюю (рис. 28.18). На предранней стадии начинается синтез РНК с двух промоторов - PL и PR. Один из образующихся при этом транскриптов служит матрицей для синтеза белка N, которому принадлежит важнейшая регуляторная роль. В отсутствие белка N предранние транскрипты заканчиваются на одном из двух участков терминации. Белок N препятствует терминированию транскрипции в этих участках и обеспечивает таким образом дальнейшее выражение генов фага X. Белок N включает раннюю стадию. В это время синтезируются белки, необходимые для репликации ДНК фага и рекомбинации. Кроме того, на ранней стадии транскрибируется ген Q. Белок Q - еще один важный регуляторный элемент выражения генов фага ![]() . Он необходим для перехода в позднюю стадию. На поздней стадии транскрибируются гены, необходимые для образования головки и отростка фага и для лизиса клетки-хозяина. Белок Q, подобно белку N, подавляет терминацию транскрипции. Короче говоря, последовательная регуляция литического развития осуществляется двумя белками - положительными регуляторами, кодируемыми генами N и Q. Их действие заключается в том, что они позволяют РНК-полимеразе продолжать транскрипцию, проскочив несколько участков терминации.
. Он необходим для перехода в позднюю стадию. На поздней стадии транскрибируются гены, необходимые для образования головки и отростка фага и для лизиса клетки-хозяина. Белок Q, подобно белку N, подавляет терминацию транскрипции. Короче говоря, последовательная регуляция литического развития осуществляется двумя белками - положительными регуляторами, кодируемыми генами N и Q. Их действие заключается в том, что они позволяют РНК-полимеразе продолжать транскрипцию, проскочив несколько участков терминации.
Рис. 28.18. Три стадии транскрипции при литическом цикле развития фага ![]() . Белок N образуется на предранней стадии и активирует раннюю стадию. Тогда в свою очередь синтезируется белок Q, который активирует позднюю стадию
. Белок N образуется на предранней стадии и активирует раннюю стадию. Тогда в свою очередь синтезируется белок Q, который активирует позднюю стадию
В лизогенном цикле различают три стадии: установление лизогенного состояния, его поддержание и выход из лизогенного состояния. Для установления состояния профага необходимо, чтобы вирусная ДНК интегрировалась с ДНК клетки-хозяина и литические функции вируса были инактивированы. Эти процессы протекают очень сложно, и до конца они не изучены. Поддержание состояния профага, наоборот, сравнительно несложный процесс. А. Дейл Кейзер (A. Dale Kaiser) показал, что из всех генов профага экспрессируется только ген сI. Этот ген кодирует X-репрессор, который связывается с двумя операторными участками OL и OR (рис. 28.19). Связывание ![]() -репрессора с OL непосредственно препятствует транскрипции ранних генов в левую сторону. В частности, не синтезируется белок N, и поэтому литический путь оказывается закрытым. Связываясь с OR,
-репрессора с OL непосредственно препятствует транскрипции ранних генов в левую сторону. В частности, не синтезируется белок N, и поэтому литический путь оказывается закрытым. Связываясь с OR, ![]() -репрессор препятствует выражению генов cro и Q в правую сторону. Таким образом, когда
-репрессор препятствует выражению генов cro и Q в правую сторону. Таким образом, когда ![]() -репрессор связан с OL и OR, весь геном фага молчит, кроме гена d, кодирующего Х-репрессор. Как будет указано чуть ниже, Х!!!!!!!-репрессор сам контролирует работу гена cI, регулируя таким путем собственную концентрацию. Инактивация
-репрессор связан с OL и OR, весь геном фага молчит, кроме гена d, кодирующего Х-репрессор. Как будет указано чуть ниже, Х!!!!!!!-репрессор сам контролирует работу гена cI, регулируя таким путем собственную концентрацию. Инактивация ![]() -репрессора обеспечивает возможность транскрипции генов, участвующих в литическом процессе. Профаг исключается из хромосомы клетки- хозяина, и литические функции экспрессируются.
-репрессора обеспечивает возможность транскрипции генов, участвующих в литическом процессе. Профаг исключается из хромосомы клетки- хозяина, и литические функции экспрессируются.
28.12. Два оператора фага лямбда содержат ряд участков связывания репрессора
-Репрессор был выделен и подробно изучен Марком Пташне (Mark Ptashne). Мономере массой 26 кДа находится в равновесии с олигомерами. С ДНК связываются именно олигомеры. Два операторных участка - ОL и ОR - узнаются одним и тем же репрессором. Ген с/, кодирующий этот репрессор, расположен между ОL и ОR
(рис. 28.19). Каждый их этих операторов содержит три участка связывания ![]() -репрессора. Расщепление с помощью нуклеаз показало, что участки связывания представляют собой последовательность из 17 пар оснований и отделены друг от друга АТ-богатыми участками длиной от 3 до 7 пар оснований. Последовательности оснований всех этих участков связывания
-репрессора. Расщепление с помощью нуклеаз показало, что участки связывания представляют собой последовательность из 17 пар оснований и отделены друг от друга АТ-богатыми участками длиной от 3 до 7 пар оснований. Последовательности оснований всех этих участков связывания ![]() -репрессора сходны, но не идентичны. Узнаваемая последовательность - 5'-ТAТСAССGС-3' или что-нибудь в этом роде. Как и lас-оператор, эти операторные участки обладают частичной симметрией второго порядка.
-репрессора сходны, но не идентичны. Узнаваемая последовательность - 5'-ТAТСAССGС-3' или что-нибудь в этом роде. Как и lас-оператор, эти операторные участки обладают частичной симметрией второго порядка.
Рис. 28.19. Схематическое изображение операторных участков OL и OR и прилегающих генов. Самое высокое сродство к Х-репрессору имеют ОL1 и ОR1. сI - ген-репрессор. Левый транскрипт начинается с гена N, правый - с гена cго
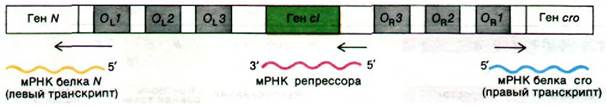
Самые сильные места связывания репрессора в операторах OL и ORрасположены ближе всего к началу первого структурного гена оперона. Промоторный участок гена Nрасположен в пределах OL а промотор гена cro - внутри OR. Как и в лактозном, и в арабинозном оперонах, связывание репрессора с этими операторами препятствует связыванию РНК-полимеразы с соответствующим промотором, и, следовательно, транскрипция не начинается. Связывание ![]() -репрессора с двумя участками в OL и OR более эффективно блокирует промотор, чем связывание только с одним участком.
-репрессора с двумя участками в OL и OR более эффективно блокирует промотор, чем связывание только с одним участком.
28.13.  -Репрессор регулирует собственный синтез
-Репрессор регулирует собственный синтез
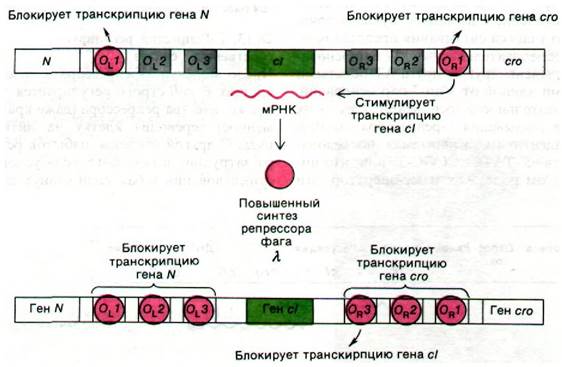
Число молекул Х-репрессора в лизогенных клетках Е. coli строго регулируется. Снижение количества репрессора (даже кратковременное) переводит клетку на литический путь. С другой стороны, избыток репрессора затруднит выход фага, если условия его существования в бактерии станут неблагоприятными. Как регулируется концентрация Х-репрессора? Проведенные сравнительно недавно исследования показали, что это репрессор регулирует собственный синтез. Для этого Х-репрессор связывается с ОR3 операторным участком, локализованным ближе всего к гену еI, и выключает транскрипцию этого гена (рис. 28.20). Связывание репрессора с ОR1, напротив, усиливает транскрипцию гена cI. Напомним, что сродство ![]() -репрессора к ОR1 выше, чем к OR3. Таким образом, транскрипция гена cI усиливается
-репрессора к ОR1 выше, чем к OR3. Таким образом, транскрипция гена cI усиливается
при низкой концентрации Х-репрессора и подавляется при высокой концентрации того же белка. Другими словами, выражение гена сI - саморегулирующаяся система.
Описанная регуляция по принципу обратной связи стремится поддерживать концентрацию репрессора на таком уровне, чтобы остальной геном фага не экспрессировался. Как же тогда фаг может выйти из состояния лизогении? Литический цикл запускается при снижении числа молекул Х-репрессора; содержание Х-репрессора должно уменьшаться до такого уровня, которого достаточно только для транскрипции гена cго. Новообразованный белок cго связывается с ОR3 и подавляет транскрипцию гена cI.
Рис. 28.20. Саморегуляция концентрации ![]() -репрессора. А - когда репрессора мало, он связывается с 0R1 и стимулирует транскрипцию гена сI. Б - по мере увеличения концентрации
-репрессора. А - когда репрессора мало, он связывается с 0R1 и стимулирует транскрипцию гена сI. Б - по мере увеличения концентрации ![]() -репрессора он связывается с ОR3 и ингибирует дальнейшую транскрипцию гена cI
-репрессора он связывается с ОR3 и ингибирует дальнейшую транскрипцию гена cI
Самое главное, что OR3 имеет более высокое сродство к белку его, чем 0R1. Таким образом, небольшое количество белка его подавляет синтез Х-репрессора, не выключая синтеза самого белка его. Вследствие этого Х-репрессор уже не может руководить ходом событий. С этого момента необратимо запускается цепь реакций, ведущих к лизису. Таким образом, тонкое взаимодействие всего нескольких белков и операторных участков определяет путь развития фага. Было бы интересно выяснить, имеют ли некоторые регуляторные системы, такие, как множественные операторные участки с различным сродством к белкам, более общее значение в регуляции развития прокариотических клеток.