БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 35. ДЕЙСТВИЕ ГОРМОНОВ
Гормоны - это химические посредники, координирующие активность различных клеток многоклеточного организма. Термин «гормон» был впервые использован в 1904 г. Уильямом Бэйлиссом и Эрнстом Старлингом (William Bayliss, Ernest Starling) при описании действия секретина - вещества, секретируемого двенадцатиперстной кишкой и стимулирующего выделение сока поджелудочной железы. Из этой работы вытекала очень плодотворная концепция, а именно, что 1) гормоны - это молекулы, синтезируемые специфическими тканями (железами); 2) гормоны секретируются непосредственно в кровь, которая и доставляет их к месту действия; 3) гормоны специфическим образом меняют активность определенных чувствительных тканей (органов-мишеней или клеток-мишеней). По своей химической природе гормоны очень разнообразны. Некоторые, например, адреналин и тироксин, представляют собой небольшие молекулы, производные аминокислот. Другие, в частности окситоцин, инсулин и тиреотропин (тиреотропный гормон), полипептиды или белки. Третья группа гормонов - стероиды, производные холестерола. В настоящее время выяснен молекулярный механизм действия ряда гормонов. Установлено, что гормоны оказывают специфическое действие тремя путями: 1) воздействуя на скорость синтеза ферментов и других белков; 2) изменяя скорость ферментативного катализа; 3) изменяя проницаемость клеточных мембран. Любопытно отметить, что ни один гормон не является ферментом или коферментом. Напротив, действие гормонов сводится к регуляции уже существующих процессов.
35.1. Открытие циклического АМР - посредника в действии многих гормонов
Важнейшее достижение в изучении механизма действия гормонов связано с именем Эрла Сазерленда (Earl Sutherland). Первоначальная цель его работ, начатых в 50-х годах, состояла в том, чтобы выяснить механизм действия адреналина и глюкагона на распад гликогена и образование глюкозы в печени. Сазерленд избрал эту систему, во-первых, потому, что указанные гормоны оказывают очень значительное и воспроизводимое действие на распад гликогена. Во-вторых, этот эффект развивается в течение нескольких минут. В- третьих, срезы печени нетрудно получить в большом количестве. В-четвертых, биохимия распада гликогена уже была достаточно хорошо изучена (гл. 16). Фактически Сазерленд начал свои исследования механизма действия адреналина и глюкагона в лаборатории Карла и Герти Кори (Carl Cori, Gerty Cori).
На начальном этапе работы он стремился выявить ту ферментативную реакцию в процессе превращения гликогена в глюкозу, которую усиливали эти гормоны. Для этого срезы печени инкубировали в присутствии 32Piи определяли, в какие промежуточные соединения включалась метка. Оказалось, что ферментом, лимитирующим скорость процесса расщепления гликогена, была фосфорилаза, а не фосфоглюкомутаза или глюкозо-6-фосфатаза. Более того, адреналин и глюкагон увеличивали активность фосфорилазы. Однако механизм активации был неясен. Далее Сазерленд обнаружил фермент, катализировавший инактивацию активной фосфорилазы. Этот инактивирующий фермент оказался фосфатазой; напрашивалось предположение, что активация фосфорилазы обусловлена ее фосфорилированием. Действительно, при инкубации срезов печени с 32Рi обнаружилось, что скорость включения метки в фосфорилазу возрастала в присутствии адреналина и глюкагона, причем это возрастание было прямо пропорционально действию гормонов на распад гликогена.
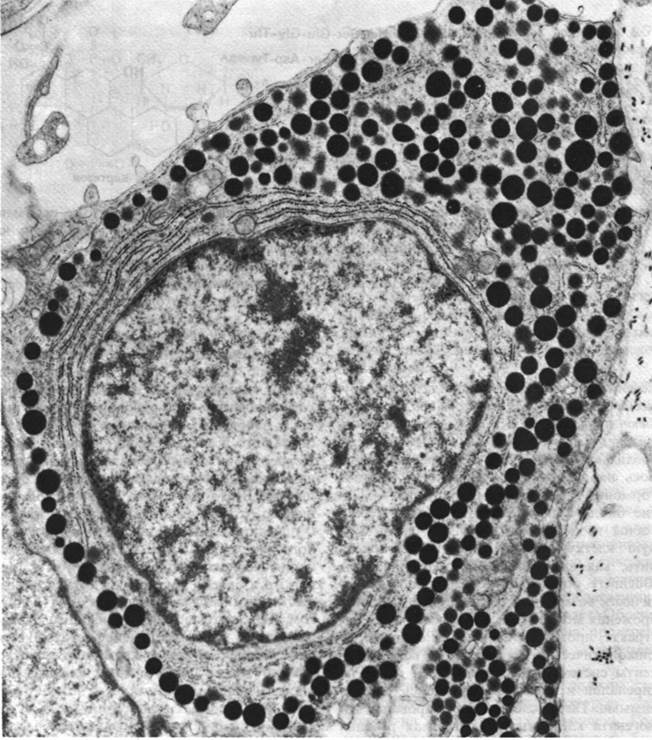
Рис. 35.1. Электронная микрофотография соматотропной клетки гипофиза. Гормон роста (соматотропин) накапливается в электронноплотных гранулах, четко видных на микрофотографии
Таким образом, в результате этих исследований и было установлено, что фосфорилаза активируется при фосфорилировании и инактивируется при дефосфорилировании. Это был первый пример регуляции активности фермента с помощью механизма ковалентной модификации.
Далее приступили к анализу активации фосфорилазы гормонами в препарате разрушенных клеток печени. Поразительным образом добавление адреналина и глюкагона приводило, как и в опытах со срезами печени, к активации фосфорилазы. Обнаружение того факта, что гормональный эффект проявляется и в бесклеточном гомогенате, явилось новой вехой в развитии биохимии. Дело в том, что ранее не удавалось наблюдать специфического действия гормонов в бесклеточных системах, и многие биологи полагали, что гормоны способны воздействовать только на интактную клетку-мишень. Любопытно вспомнить, как за полвека до этого Бухнеры (Buchner) опровергли совершенно аналогичные воззрения, доказав, что процесс брожения может идти и в бесклеточном экстракте дрожжей. Однако в отличие от гликолитических ферментов не все компоненты системы, обеспечивающей ответ на адреналин и глюкагон, оказались растворимыми. Так, после центрифугирования гомогената клеток печени ответная реакция на добавление гормонов исчезала. Следовательно, какая-то существенная часть системы гормонального ответа была локализована во фракции мембран. Действительно, гормональный ответ можно было восстановить, добавив к надосадочной жидкости фракцию субклеточных частиц.
Рис. 35.2. Примеры трех химически различных классов гормонов: А - адреналин, производное аминокислоты; Б - глюкагон, полипептид; В - кортизол, стероид

Роль субклеточных частиц была выявлена следующим образом. При инкубации этой фракции с адреналином и глюкагоном образовывался некий термостабильный фактор. Добавление этого фактора к надосадочной фракции приводило к активации фосфорилазы. Другими словами, гормональный ответ удалось разделить на два этапа: взаимодействие гормона с мембранной фракцией, приводящее к образованию термостабильного фактора, и действие этого фактора на надосадочную фракцию, выражающееся в активации фосфорилазы. Следующая задача состояла в том, чтобы идентифицировать термостабильный фактор, полученный в очень малом количестве. По данным химического анализа это был аденинрибонуклеотид, но с необычными свойствами. Сазерленд описал его в письме Леону Хеппелю (Leon Heppel), к которому обратился в надежде на помощь в изучении структуры этого вещества. В это же время Хеппель получил письмо от Дэвида Липкина (David Lipkin) с описанием нового нуклеотида, полученного путем обработки АТР гидрооксидом бария. Хеппель пришел к выводу, что Липкин и Сазерленд изучают одно и то же вещество, и помог им связаться друг с другом. Действительно, оба ученых исследовали одно и то же соединение, оказавшееся аденозин-3',5'-монофосфатом, или, как его обычно называют теперь, циклическим AMP (cAMP). Эта случайная встреча двух ученых принесла еще одну пользу: сразу возникла возможность получения больших количеств сАМР для биохимических исследований. Более того, в результате лабораторного синтеза сАМР из АТР и гидрооксида, бария появилась возможность выдвинуть предположение о вероятном пути биосинтеза этого соединения.
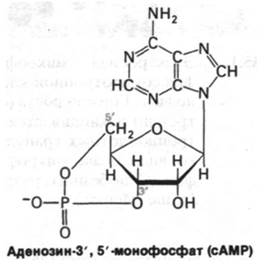
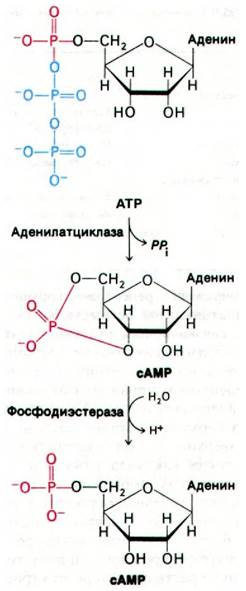
Рис. 35.3. Ферментативный синтез и распад сАМР