Біофізика і біомеханіка - В. С. Антонюк - 2012
Розділ 4. БІОФІЗИКА СКЛАДНИХ БІОЛОГІЧНИХ СИСТЕМ
4.2.Особливості керування руховою функцією організму
Унаслідок аналізу та оцінювання у нервовій системі тієї інформації, яку організм отримує через свої сенсорні системи, виникає певна діяльність організму. Як закономірна відповідь організму на зовнішній вплив через посередництво нервової системи вона виявляється як рефлекторна реакція. Основним компонентом такої реакції є скорочення м’язів, за допомогою яких організм може наблизитися до подразника, якщо він становить для нього інтерес, віддалитися від подразнення, що може спричинити пошкодження, тощо.
Кожний м’яз, якщо його розглядати відокремлено від нервових центрів, поводить себе як пасивна еластична структура, здатна до пасивного розтягування під впливом зовнішньої сили. Але коли м’яз включений у систему аферентних та еферентних зв’язків з цими центрами, то його біофізичні характеристики істотно змінюються і він набуває властивостей системи, що саморегулюється та змінює свій стан залежно від ступеня розтягнення.
Ця здатність забезпечується власними рефлексами м’язів, рецепторні структури яких містяться у самих м’язах і які є сенсорами (датчиками) фізичних характеристик їх стану (довжини та напруження). Наявність таких механізмів дає змогу м’язам за будь-якого положення чи руху кінцівок і тулуба зберігати певний вихідний фізичний стан, від якого надалі можуть перебігати ті чи інші рухові реакції.
Рецепторний апарат м’язів. Сенсорами стану м'яза є два типи м’язових рецепторів: м’язові веретена та сухожилкові органи. М’язові веретена являють собою більш тонкі й короткі м’язові волокна, що зібрані у веретеноподібні пучки та вкриті зовні сполучнотканинною капсулою.
У рецепторах м’язових волокон закладено механізм регуляції їхньої чутливості, який керується сигналами з центральної системи. Така система може компенсувати ті зміни в чутливості м’язового рецепторного апарата, які виникають у разі зміни фізичного стану м’яза - його розтягнення чи скорочення залежно від положення місць його прикріплення до кісткової системи і відповідно розтягнення чи скорочення інтрафузальних м’язових волокон. Точність такого регулювання посилюється тією обставиною, що сигнали з боку гамма-волокон можуть бути двох типів - динамічними та статичними. Динамічні сигнали змінюють чутливість м’язових рецепторів до швидкості розтягнення м’яза, але незначно впливають на їхню стаціонарну чутливість, статичні ж ефективно змінюють частоту аферентних сигналів саме у процесі постійного розтягування м’яза. Таким чином, регулюванню підлягають як пропорційні, так і диференційні характеристики функціонування м’язових рецепторів.
Механізми саморегуляції стану м’яза. Складний рецепторний апарат є основою саморегуляторного механізму, дуже важливого для успішного функціонування скелетної мускулатури. Цей механізм містить поряд із системою рецепторів та відповідних аферентних волокон відповідні нейронні структури спинного мозку, до яких надходять аферентні сигнали і в яких вони переключаються на рухові (моторні) нейрони, що надсилають еферентні (рухові) команди до тих же м’язів.
М’яз скорочується, протидіючи тим самим його розтягненню. Навпаки, у разі скорочення м’яза синаптична дія на рухові нейрони послаблюється і відповідно зменшується сила скорочення м’яза.
Мембрана м’язових клітин - сарколема, як і мембрана нервових клітин, електрозбудлива і здатна проводити потенціал дії. Ці процеси в м’язових клітинах відбуваються за тим же принципом, що і в нервових. Потенціал спокою (V0) м’язового волокна становить близько мінус 90 мВ, що нижче від такого ж у нервовому волокні (-70 мВ), а критична деполяризація (Vк), з досягненням якої виникає потенціал дії, має однакове значення з нервовими волокнами. Тому збудливість м’язового волокна дещо нижча, ніж нервового.
Відповіддю м’язового волокна на збудження є скорочення, яке виконує скорочувальний апарат клітини - міофібріли. Вони складаються з двох видів ниток: товстих - міозинових і тонких - актинових.
Кожна товста нитка оточена шістьма тонкими, а кожна тонка - трьома товстими. Таким чином, у поперечному зрізі м’язове волокно має правильну гексагональну структуру.
У разі скорочення м’яза довжина актинових і міозинових філаментів не змінюється. Відбувається лише їх зсув відносно один одного: тонкі нитки засовуються в проміжки між товстими.
Для того щоб актин і міозин могли взаємодіяти, потрібні іони кальцію. Якщо м’яз розслаблюється, активізується робота кальцієвого насоса, що знижує концентрацію Са2+ в цитоплазмі; отже, зв язки між тонкими і товстими нитками вже не можуть утворюватися. У цих умовах під час розтягування м’яза нитки безперешкодно ковзають одна відносно одної. Проте така розтяжність можлива тільки за наявності АТФ. Якщо в клітині немає АТФ, то актин-міозиновий комплекс не може розірватися. Нитки залишаються жорстко зчепленими між собою. Це явище спостерігається у разі трупного окоченіння.
Є два режими скорочення м’яза: ізотопічне (змінюється довжина волокна, а напруження залишається незмінним) й ізометричне (кінці м’яза нерухомо закріплені, унаслідок чого змінюється не довжина, а напруження).
Важливими характеристиками роботи м’яза є сила і швидкість скорочення.
Рівняння Хілла, що зв’язує між собою силу та швидкість скорочення м’яза, має вигляд:![]() де υ - швидкість скорочення м’яза; Р - м’язова сила або прикладене до неї навантаження (зовнішня сила, яка діє на м’яз); υmaχ - максимальна швидкість скорочення м’яза (υ = υmaχ, якщо P = 0); Р0 - сила, яка розвивається м’язом в ізометричному режимі скорочення, тобто з таким навантаженням, за якого довжина м’яза не змінюється (Р = Р0, якщо υ = 0); а і b - константи. Якщо м’яз скорочується за час t, він виконує роботу А:
де υ - швидкість скорочення м’яза; Р - м’язова сила або прикладене до неї навантаження (зовнішня сила, яка діє на м’яз); υmaχ - максимальна швидкість скорочення м’яза (υ = υmaχ, якщо P = 0); Р0 - сила, яка розвивається м’язом в ізометричному режимі скорочення, тобто з таким навантаженням, за якого довжина м’яза не змінюється (Р = Р0, якщо υ = 0); а і b - константи. Якщо м’яз скорочується за час t, він виконує роботу А:
![]()
У процесі скорочення м’яза виділяється деяка кількість теплоти Q. Цю величину називають теплопродукцією. Як показав Хілл, теплопродукція залежить тільки від зміни довжини х м’яза і не залежить від навантаження Р:Q = ах.
Загальна потужність Nзаг, розвинута м’язом, визначається швидкостями виконання м’язом роботи та виділення теплоти:
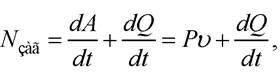
де Ρυ = Nкор - корисна потужність. Коефіцієнт корисної дії м’яза зберігає стале значення (близько 40 % ) у діапазоні значень сили від 0,2 Р0 до 0,8 Р0.
Таким чином, за принципом сервомеханізму відбувається автоматична компенсація змін навантаження на м’яз, напрямлена на збереження його постійної довжини (розтягнення).
Оскільки сухожилкові рецептори являють собою датчики напруження м’яза, то їх активація буде напрямлена на зниження цього напруження за м’язового скорочення. Навпаки, зі зниженням м’язового
тонусу будуть розгальмовуватися відповідні рухові нейрони і підвищуватиметься їх активність. Таким чином, створюється ще один сервомеханізм, напрямлений на компенсацію змін напруження м’яза та збереження його оптимального рівня.
Жорсткість м’язів має велике значення, оскільки вона визначає їхню поведінку у разі невеликих змін їх довжини, що зазвичай відбувається за умов фізіологічної діяльності. Так, якщо підтримується поза тіла, вона забезпечує стабілізацію взаємного положення окремих частин тіла та кінцівок за різних відхилювальних впливів. Таке регулювання стану м’язів позначається як підтримання їхньої короткодіапазонної жорсткості.
Вивчення опорно-рухової системи людини має двояке завдання. По-перше, знання динаміки опорно-рухової системи проливає світло на закономірності руху живого організму в просторі та часі, по-друге, допомагає запозичувати для техніки, наприклад для роботобудування, багато корисного з кінематики та динаміки руху тварин і людей.
Опорно-руховий механізм людини є дуже складною системою з багатьма степенями вільності, що визначаються кількістю незалежних величин, які мають задаватися для визначення положення тіла (або системи тіл). Отже, вільне абсолютно тверде тіло має шість степенів вільності: три взаємно перпендикулярні переміщення і три обертання навколо взаємно перпендикулярних осей.
Скелет дорослої людини складається з 206 кісток, багато з яких сполучені так, що допускають взаємний рух (наприклад, кістки руки або кістки ноги). У скелеті приблизно 150 з’єднань, з яких майже половина - суглоби. Розглянемо спрощену схему руки. Як показано на рис. 4.9, плечовий суглоб можна умовно зобразити шарніром, що має три степені вільності (три незалежні обертання).
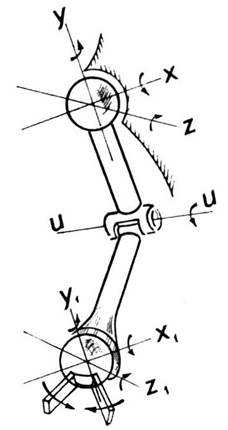
Рис. 4.9. Кінематична схема плечовoго суглоба
Ліктьовий суглоб допускає лише згинання та розгинання руки (ще одне обертання), а суглоб перед кистю також можна змоделювати шарніром (ще три обертання). Тепер якщо вважати кисть «захопленням», що виконує дії «узяти - відпустити», тоді вся система матиме вісім степенів вільності. Таку модель можна використовувати для конструювання маніпулятора.
Проте математична модель ускладнюється зі збільшенням кількості степенів вільності. Кількість степенів вільності, наприклад людського тіла, приблизно дорівнює 240.
До динамічних моделей опорно-рухового апарата можна успішно застосовувати методи класичної механіки. Скелет людини і тварини є своєрідним каркасом для тіла. Він приводиться в рух скелетними м’язами, кожний з яких прикріпляється до двох різних кісток (рис. 4.9). Зі скороченням м’яза її довжина зменшується, унаслідок чого змінюється кут між відповідними кістками.
Як ілюстрацію діяльності м’язів розглянемо одне з простих завдань біомеханіки - утримання рукою вантажу (рис. 4.10).
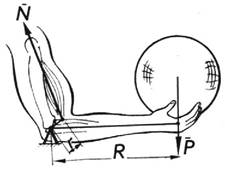
Рис. 4.10. Кінематична схема утримання рукою вантажу
У цьому завданні, вперше розглянутому ще Леонардо да Вінчі, за відомими довжинами кісток плеча і передпліччя, величиною вантажу і положенням точок кріплення м’яза треба знайти силу, що розвивається м’язом. Згідно із сучасними поняттями - це елементарна задача, проте досить важливим є результат її розв’язання: кінематична схема плечевого суглоба може бути описана як важель другого роду, зусилля м’язів якого у стільки разів більше від величини навантаження, у скільки разів точка закріплення м’язів ближча до ліктьового суглоба, ніж вантаж, який утримується. Це можна подати таким математичним виразом: Nr = PR, де N - зусилля, яке розвиває м’яз; r - відстань від точки кріплення м’яза до кістки передпліччя до ліктьового суглоба; Р - вага вантажу; R - відстань вантажу від ліктьового суглоба (рис. 4.11).
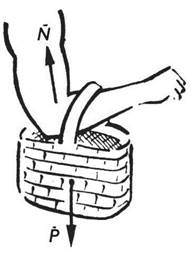
Рис. 4.11. Фізіологічний приклад важеля другого роду
Силу м’язу, яка припадає на 1 см її поперечного перерізу, називають абсолютною м’язовою силою. Для людини вона становить 50.. .100 Н.
Як приклад важеля першого роду можна навести утримання голови шийними м’язами (так, у знесиленої людини голова звішується на груди). Для аналізу можна знову застосувати математичний запис Nr = PR, але тепер важелі розміщені по різні боки опори. Тут N - зусилля, передаване від м’яза, Р - сила тяжіння, прикладена в центрі тяжкості голови; r і R - відповідні важелі.
Сила тяжіння як результат наявності гравітації безперервно притискає людину до Землі, сили пружності підтримують її на поверхні, а тертя дозволяє здійснювати переміщення. Сили неживої природи можна долати за рахунок сили м’язів, яка підкорюється велінням мозку без механічних посередників.
М’яз - це чудовий природний механізм. Якщо його умовно назвати машиною, то перш за все привертає увагу її економічність. М’язи черепахи, наприклад, мають ККД близько 80 %, тоді як ККД кращої парової турбіни - не більше 40 % [108]. Це є справедливим не тільки для опорно-рухової системи. Аналогічна ситуація спостерігається у разі спроби розгризти зубами горіх. Зробити це намагаються корінними зубами, які ближче до місця закріплення жувальних м’язів. Очевидно, що в цьому випадку зусилля з боку зубів на шкаралупу горіха буде найбільшим. От чому і собака, що гризе кістку, прагне скористатися найдалі розміщеними зубами. У процесі природного відбору корінні зуби спеціально пристосувалися для такої діяльності.
Кожна людина може встати навшпиньки на одній нозі і навіть зі значним вантажем. Литковий м’яз здатен підняти близько 100 кг вантажу. Якщо ж урахувати, що стопа є важелем першого роду і литковий м’яз прикріплений до короткого плеча цього важеля, то можна встановити, що литковий м’яз навантажений так, немовби до його нижнього кінця прикріпили вантаж масою 1 т (рис. 4.12). Зусилля для опорного стрибка на ногу спортсмена таке, неначе спортсмен раптом почав важити тонну.
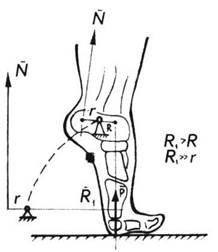
Рис. 4.12. Фізіологічний приклад важеля першого роду
Проте, як відомо з фізіології [92], людина по своїй волі не може надати м’язу максимального скорочення. У разі порушення регуляції з боку нервової системи м’яз може розвинути свою повну, максимальну силу, і вона в цій ситуації здатна відірвати шматочок кістки, до якої прикріплена. Як відомо, потопаючих зазвичай рятують, витягуючи їх за волосся. Це тому, що захоплення рукою може бути згубним для обох: розтиснути руку потопаючого майже неможливо.
Основу м’язової діяльності становлять складні хімічні перетворення усередині клітин, що приводять до механічного ефекту - скорочення м’яза. Ланцюг процесів, що перебігають у м’язі, який скорочується, складний і ще далеко не вивчений у всіх тонкощах.