ФІЗІОЛОГІЯ ТА БІОХІМІЯ РОСЛИН
Опорний конспект лекцій
3. СИСТЕМИ РЕГУЛЯЦІЇ ТА ІНТЕГРАЦІЇ У РОСЛИН
Міжклітинні системи регуляції
Трофічна регуляція
Важливою складовою частиною забезпечення цілісності рослинного організму, регуляції його гомеостазу є трофічні взаємодії між різними частинами рослини. Взаємодія з допомогою поживних речовин - найпростіший взаємозв'язок між клітинами, тканинами та органами. Як відомо, система живлення у рослин двокомпонентна: повітряне живлення здійснюється в основному листками, а поглинання води і мінеральних речовин - корінням. Рослинний корінь та інші гетеротрофні органи залежать від надходження речовин-асимілятів, котрі утворилися в листках у процесі фотосинтезу (цукор, вітаміни, нікотин і т.п.). У свою чергу надземні частини мають потребу в мінеральних речовинах та воді, які поглинаються з ґрунту коренем.
Зсуви у вмісті різних елементів живлення впливають на обмін речовин, фізіологічні та морфогенетичні процеси у рослин. Однак трофічна регуляція носить швидше кількісний, ніж якісний, характер. При голодуванні у рослин, як правило, розвиток продовжується відповідно до внутрішніх закономірностей, але у них формуються органи меншого розміру і зменшується кількість листків, плодів, насіння. При цьому кінцева величина насіння залишається в нормі.
Гормональна система регуляції
Фітогормони - сполуки, із допомогою яких здійснюється взаємодія клітин, тканин та органів і котрі в незначних кількостях необхідні для запуску та регуляції фізіологічних і морфогенетичних програм.
Гормони рослин - порівняно низькомолекулярні органічні речовини (Mr 28-346). Вони утворюються в різних тканинах та органах і діють у дуже низьких концентраціях (порядку 10-12-10-5 моль/л). Уперше їх виявили Ч. Дарвін (1881) та Г. Фіттінг (1910).
Великий вклад у розвиток гормонального напрямку щодо фізіології рослин вніс український фітофізіолог М.Г. Холодний. 1918 року він висловлює думку про те, що верхівки кореня, стебла, молоді листки є органами внутрішньої секреції, які виділяють до зони росту речовини гормонального характеру. Згодом М.Г. Холодний створив фітогормональну теорію тропізмів (теорія Холодного-Вента). Досліджуючи причини рухів, кореляцій, переходу рослин від спокою до активного росту він дійшов висновку, що каталізаторами та регуляторами ростових і морфогенетичних процесів виступають гормони, вітаміни, ферменти. У 1939 році вийшла монографія вченого «Фітогормони», яка мала надзвичайно глибокий вплив на подальший розвиток досліджень із фітогормонології, фізіології росту. Надалі ці напрямки розвивали К. Ситник, Л. Мусатенко, Т. Богданова (1978). Слід зауважити, що фітогормональна система менш спеціалізована порівняно з ендокринною системою тваринних організмів. У рослині для запуску чи виключення будь-яких морфогенетичних та фізіологічних програм використовуються ті самі гормони, тільки в різних співвідношеннях.
Встановлено, що у вищих рослин міститься кілька важливих класів гормонів: індоліл-3-оцтова кислота (ауксин) - ІОК, цитокініни (ЦК), гібереліни (ГА), етилен (Ет), абсцизини (АБК). Останнім часом до них зараховують і брасини (брасиностероїди), жасмонову кислоту, а також фенольні інгібітори росту (ФІ) негормональної природи.
Умовно перші три групи (ауксини, гібереліни та цитокініни) і частково брасини можна вважати речовинами стимулюючої дії, тоді як абсцизову кислоту, етилен і жасмонову кислоту - інгібіторами. Гібереліни та цитокініни трапляються у вигляді споріднених, подібно діючих груп молекул, тоді як інші - у природі представлені лише однією сполукою. В цілому, ріст і розвиток будь-якої тканини зумовлений наявністю всіх цих речовин, які синтезуються в різних частинах рослини.
Ауксини (ІОК)
Перші з фітогормонів, які були досліджені, - це ауксини - речовини індольної природи. Природний ауксин було ідентифіковано в 1934 р. у лабораторії Ф. Кегля як індоліл-3-оцтову кислоту з верхівок колеоптилей злаків. Тепер відомо багато синтетичних препаратів, які стимулюють ріст подібно до індолілоцтової кислоти (індолілпіровиноградна, хлорфеноксіоцтова, 2,4-дихлорфеноксіоцтова кислоти та ін.). Вони дешевші і часто активніші, адже в рослин немає ферментів для їх розщеплення.
ІОК широко поширена в рослинному світі, у представників прокаріот і грибів. У вищих рослин її найбільше у бруньках та листках, що розвиваються, в активному камбії, в насінні, що формується, у пилку. В цілій рослині ауксину найбільше на верхівці головного пагону.
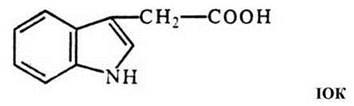
Попередники ауксину - триптофан і триптамін. ІОК може також вивільнятися зі зв'язаного стану (із глікозидів, комплексів із білками чи амінокислотами). Різні похідні індолу є попередниками ауксину, вони можуть у рослині перетворюватися на ІОК.
Транспорт ІОК паренхімними та камбіальними тканинами йде полярно із швидкістю 10-15 мм/год від верхівки до кореня. У стеблі рух відбувається провідними пучками. Механізм полярного транспорту такий. В апікальний кінець клітини гормон проникає пасивно разом з іонами Н+; а в базальному кінці активно секретується через клітинну мембрану. В листках ауксин пересувається по флоемі. Цей рух не є полярним, він збігається з транспортуванням вуглеводів.
Інгібітори транспорту → нафтілфталамінова кислота, морфактин.
Фізіологічна дія ауксину. Ауксин різноманітно впливає на рослину, залежно від етапу онтогенезу, виду рослин і, особливо, типу тканини. У високих концентраціях він токсичний. Гербіцид 2,4-Д - синтетичний ауксин для боротьби з бур'янами.
Найбільш виражений ефект ауксину виявляється в стимуляції росту розтягуванням. Ауксин активує поділ і розтягування клітин, необхідний для формування провідних пучків і коренів, сприяє розростанню опліддя. Перше практичне застосування ауксину базувалося на його здатності стимулювати утворення корінців у живців.
Тканини, багаті на ауксин, володіють атрагуючою властивістю (притягування поживних речовин). У багатьох випадках обробка гормоном затримує процеси старіння тканин та органів. Ауксин зумовлює явне апікальне домінування. Він здатний затримати розпускання бічних бруньок та опадання листків і плодів. Застосовуючи ауксини у виробництві, можна регулювати листопад, опадання плодів, квіток. Ауксини впливають на в'язкість протоплазми, посилюють поглинання води багатьма тканинами (наприклад, паренхімою бульб картоплі). Гормон відіграє головну роль у ростових рухах - тропізмах і настіях.
Молекулярні механізми дії. Проникаючи до клітин, ІОК зв'язується зі спеціальними рецепторами, діючи на функціональну активність мембран, полірибосом і роботу ядерного апарату. У плазмалемі ауксин індукує роботу Н+- помпи, в результаті чого матрикс клітинних стінок закислюється. Це призводить до посилення активності кислих гідролаз і розм'якшення клітинних стінок, що є необхідною умовою для росту клітин розтягуванням. Ймовірно, існують два типи впливу ауксину на фазу розтягування. Перший, короткочасний, зумовлюється «кислим ростом», а потім спостерігається довготривалий вплив, пов'язаний з регуляцією експресії щонайменше 10-ти специфічних генів, які відповідають за процеси росту.
Майже всі тканини можуть розщеплювати ауксин із допомогою ІОК-оксидази. Фермент окиснює ІОК там, де вона більше не потрібна. Також велике значення має руйнування гормону на світлі (фотоокиснення).
Цитокініни
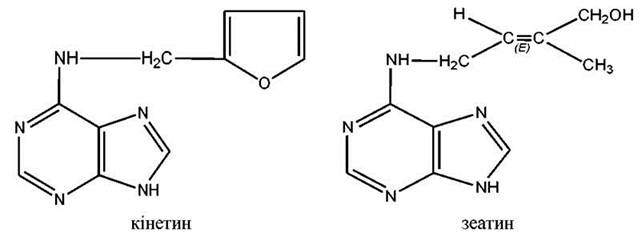
Речовини, необхідні для індукції поділу рослинних клітин, отримали назву цитокінінів. Відкриття цитокінінів пов'язане з дослідженнями культури тканин. У 50-х роках було виявлено, що клітини калусу в стерильній культурі на поживному середовищі через певний проміжок часу перестають ділитися. Згодом було виділено речовину, яка викликала поділ клітин. Її назвали кінетином. Дещо пізніше з недозрілих зернівок кукурудзи був виділений природний цитокінін - зеатин, який вважають найактивнішим природним гормоном цієї групи.
У даний час цитокініни виявлені у мікроорганізмах, водоростях, усіх вищих рослинах. Вони містяться в різних тканинах, зокрема, у ксилемному соці, верхівках коренів, проростаючому насінні, пухлинах корончатих галів, бульбочках бобових. Найбільше їх у плодах та насінні на початкових етапах формування і в меристематично активних ділянках. Цитокініни входять до складу всіх видів тРНК, але ці тРНК гормональною активністю не володіють.
Основне місце синтезу цитокінінів у вегетуючих рослинах - апікальні меристеми коренів. З коренів цитокініни транспортуються до надземних органів по ксилемі. Цитокініни для тРНК синтезуються кожною живою клітиною.
Усі природні цитокініни - похідні ізопентеніладеніну. Вони синтезуються шляхом конденсації аденозин-5-монофосфату і Δ2-ізопентенілпірофосфату. Зв'язані цитокініни у вигляді риботидів, рибозидів і глюкозидів є транспортними й запасними формами.
Фізіологічна дія. Цитокініни індукують поділ клітин, однак лише за наявності ауксину. Обробка рослин цитокінінами разом з ауксинами змушує диференційовані клітини знову перейти до ділення. В культурі калусної тканини цитокініни при підвищених концентраціях (0,5-1,0 мг/л) індукують утворення бруньок та пагонів.
Усі фізіологічні ефекти, зумовлені дією цитокінінів, умовно можна поділити на дві групи: 1) стимуляція поділу клітин та їх диференціювання, 2) затримка процесів старіння.
Вони діють як специфічні стимулятори мітозу, відіграють істотну роль у процесах диференціювання клітин та органогенезу. В основі ростових ефектів цитокінінів лежить процес прискореного клітинного поділу, пов'язаний з посиленим синтезом ДНК. Вони індукують утворення бруньок, їх розпускання, проростання насіння. Під впливом цитокінінів відбувається загальна стимуляція обміну речовин, насамперед, синтезу ДНК і білків. Цитокініни затримують старіння, тому що вони сприяють підтримці певного рівня білків та нуклеїнових кислот, а також шляхом збереження цілісності клітинних мембран.
Цитокініни необхідні для нормального розвитку листків і для підтримання їх атрагуючої властивості. Вони запобігають розпаду хлорофілу, впливають на структуру хлоропластів, підвищують інтенсивність фотосинтезу, фотофосфорилювання, активують рибулозодифосфаткарбоксилазу.
На молекулярному рівні цитокініни в комплексі з регуляторними білками посилюють активність РНК-полімерази і матричну активність хроматину; при цьому збільшується кількість полірибосом і активується синтез білка, зокрема, нітратредуктази. Є дані про дію цитокінінів на транспорт К+, Н+ і Са2+.
Гібереліни
Відкриття даної групи фітогормонів пов'язане з вивченням хвороби рису, яка виявлялась у сильному видовженні стебла й листків (Є. Куросава, 1926). Згодом була виділена речовина, яка спричиняла витягування пагонів (Т. Ябута, 1938). Її назвали гібереліном.
У даний час виявлено близько 90 гіберелінів кислої та нейтральної природи. Для них уведено шифр ГА. Згідно з цим шифром гіберелова кислота, що є третім відкритим гібереліном, позначається, як ГА3.
Гібереліни - численна група сполук класу тетрациклічних дитерпеноїдів, складної групи рослинних вторинних метаболітів, близьких до ліпідів. Це слабкі карбонові кислоти, які мають «гіберелановий» скелет.
Гібереліни виявлені у грибах, водоростях і вищих рослинах. Найбільша їх кількість міститься в недозрілому насінні. Різні етапи онтогенезу рослин можуть характеризуватися неоднаковим вмістом та набором гіберелінів. Синтезуються головним чином у листках, а також у коренях. Світло стимулює утворення гіберелінів. Транспортування відбувається пасивно з ксилемним та флоемним током.
Як і всі поліізопренові сполуки, гібереліни синтезуються з ацетил-СоА через мевалонову кислоту і геранілгераніол. Зв'язані у вигляді глікозидів, гібереліни є запасною й транспортною формами.
Фізіологічна дія. Вплив гіберелінів на рослини найхарактерніше виявляється у видовженні стебла, зокрема, у стимуляції росту стебла при обробці карликових мутантів кукурудзи. Гормони помітно посилюють витягування пагонів і в нормальних рослин (~ на 30-50%). Через це гіберелін вважають гормоном росту стебла. Місця дії гіберелінів - апікальні та інтеркалярні меристеми, де під впливом цього фітогормону активується в основному розтягування, а не поділ клітин.
Яскравий прояв дії цих гормонів - стимуляція стрілкування, а також цвітіння в рослин довгого дня в умовах короткого дня; у рослин же короткого дня гіберелін може викликати протилежну реакцію щодо цвітіння. Гібереліни стимулюють цвітіння у багатьох видів рослин. Екзогенно внесений гіберелін знімає у дворічних рослин необхідність у яровизації. Це не стосується тих дворічників, які формують стебло вже в перший рік вегетації.
Гібереліни не стимулюють ріст кореня, а в підвищених концентраціях погіршують його стан. Як правило, вони не потрібні в культурі тканини. Обробіток гіберелінами виводить насіння й бульби деяких рослин зі стану спокою. Гіберелова кислота викликає партенокарпію.
Припускають наявність двох шляхів контролю гіберелінами специфічних фізіологічних процесів: по-перше, через синтез гормону з наступною ініціацією гіберелінзалежного процесу; по-друге, в результаті взаємоперетворення великої кількості подібних за структурою, але різних за своєю активністю гіберелінів. Це дає змогу рослині легко керувати різноманітними процесами метаболізму, перетворюючи активний ГА в неактивний і навпаки.
Молекулярні механізми дії. Виявлена активуюча дія гіберелінів на синтез НК і білків. Як й інші фітогормони, у клітині гібереліни перш за все реагують із специфічним регуляторним білком. Вони активують ферменти, які відповідають за синтез фосфоліпідів мембран та утворення гранулярного ЕР і секрецію ферментів.
Попередні дані також свідчать про те, що гіберелін індукує розтягування клітин завдяки підкисленню клітинних стінок аналогічно до того, як це робить ауксин. Причому, як виявилося, чутливі до ГА клітини не реагують на ауксин.
Абсцизини
Для того, щоб вижити у змінних умовах навколишнього середовища, рослина повинна мати здатність у потрібний час гальмувати або стимулювати ті чи інші фізіологічні процеси. Згадаємо перехід рослинного організму до літнього спокою, підготовку до зими чи, навпаки, до пробудження навесні. Без сумніву, в цих процесах значну роль відіграють фітогормони, зокрема, абсцизова кислота (АБК). Відкриття АБК було пов'язане з вивченням двох явищ - опадання листків і плодів та стану спокою бруньок. У 1961 році із сухих коробочок бавовнику, а в 1963 - зі старих листків берези було виділено речовину, яка гальмувала ріст і переводила бруньки до стану спокою. Її назвали абсцизовою кислотою, а згодом визначили молекулярну будову (Окума, Корнфорт, 1963).
Абсцизова кислота є сесквітерпеном (С15Н20О4) і має таку структуру:
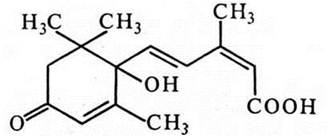
АБК виявлена у папоротеподібних, голонасінних та покритонасінних. У водоростей та мохів її функцію виконує лунуларова кислота, а у грибів та бактерій її немає. АБК багато у старих листках, зрілих плодах, бруньках та насінні у стані спокою.
Абсцизини синтезуються переважно в листках (у пластидах), у плодах, що дозрівають, та в кореневому чохлику. Синтез відбувається двома шляхами:
1) із мевалонової кислоти через ізопентилпірофосфат і геранілпірофосфат;
2) шляхом розпаду каротиноїдів через ксантоксин до АБК.
Зв'язана форма АБК - це складний ефір абсцизової кислоти з Д-глюкозою. Рух АБК в рослинах здійснюється як у базипетальному, так і в акропетальному напрямках у складі ксилемного й флоемного току.
Фізіологічна дія. У більшості випадків АБК гальмує ріст рослини. Вона виступає антагоністом ІОК, ЦК і ГА. У деяких випадках виступає як активатор (розвиток партенокарпічних плодів, видовження гіпокотилей, утворення коренів у живців і т.д.).
АБК - сильний інгібітор проростання насіння та росту бруньок і накопичується при переході рослин чи органів до стану фізіологічного спокою; призводить до утворення розмежувального шару у черешках листків та плодоніжках, прискорюючи опадання листя та плодів. АБК прискорює старіння, спричиняючи розпад НК, білків, хлорофілу.
Важливою функцією АБК є участь у механізмах стресу. Абсцизова кислота швидко нагромаджується в тканинах за дії несприятливих факторів, особливо у випадку нестачі води, викликаючи закривання продихів і зменшення транспірації. АБК бере участь у формуванні та прояві реакції кореня на гравітацію. Корінь росте вниз під впливом сили тяжіння. В механізмі явища геотропізму значну роль відіграє абсцизова кислота, яка при горизонтальному рості кореня транспортується в нижню частину, де й затримує ріст. Відповідно верхня частина росте, зумовлюючи вигин кореня вниз.
Крім того, АБК необхідна для утворення бульб, екзогенне введення її зумовлює прискорення дозрівання плодів (виноград, полуниці, томати, яблука).
Таким чином, абсцизова кислота затримує ростові процеси в усіх їх проявах. Вона контролює перехід до стану спокою, старіння листків, дозрівання плодів, бере участь у бульбоутворенні, явищах геотропізму, є гормоном стресу, інгібітором проростання насіння, бульб, росту бруньок.
Механізм дії АБК на молекулярному рівні невідомий. Вважають, що вона блокує деякі етапи процесу транскрипції, трансляції, гальмує активність Н+-помпи, змінює проникність мембран для іонів, що може спричинити різні наслідки. Важливою функцією АБК є активація експресії генів, що кодують захисні білки.
У практиці АБК використовують для обприскування плодових дерев у кінці вегетації з метою прискорення дозрівання й скорочення термінів збирання врожаю. Вміст абсцизової кислоти, особливості її деградації можуть бути важливим критерієм у селекції рослин стосовно посухостійкості. Розкриття механізмів інгібіторної дії АБК позитивно вплине на пошук екологічно безпечних, не фітотоксичних інгібіторів, ретардантів та інших синтетичних регуляторів росту у рослин.
Етилен
Газ етилен (етен, СН2=СН2) у низьких концентраціях (0,04-1,0 мкл/л) виявляє сильну морфогенну дію на рослини. Вперше фізіологічний ефект етилену описав Д. Нелюбов (1901), який виявив, що в етиольованих проростках газ викликає "потрійну реакцію" стебла: гальмування росту, потовщення і горизонтальну орієнтацію. Викликані етиленом зміни в рості не носять постійного характеру. Якщо етилен видаляли з атмосфери, то нормальний ріст відновлювався. Пізніше виявилося, що етилен прискорює дозрівання плодів (Ф. Денні, 1924). Однак майже всі вважали, що він є компонентом забруднення середовища. І лише 1934 року О. Гейну вдалося встановити, що етилен продукує сама рослина.
Етилен утворюється деякими бактеріями, грибами й вищими рослинами. Практично всі тканини покритонасінних здатні утворювати етилен. Найбільш активно це відбувається при пораненні або в стресових умовах. Найбільше його в листках, що старіють, та в плодах, котрі достигають. Виділення етилену гальмується браком кисню і може регулюватися світлом.
У вищих рослин етилен синтезується з метіоніну через S-аденозилметіонін та аміноциклопропанкарбонову кислоту. Остання є і транспортною формою та разом із етиленом, що добре розчинний у воді, переміщується з транспіраційним током. Однак вважають, що місце дії етилену наближене до того місця, де він синтезується, тому немає потреби в його транспортуванні. Концентрація етилену залежить від швидкості синтезу. В утворенні етилену бере участь низка ферментів, які активуються при дозріванні, в умовах стресу, а також ауксином і власне етиленом. Газ вільно дифундує через міжклітинний простір до оточуючого середовища.
Фізіологічна дія. Етилен гальмує подовження паростків; у дводольних зупиняє ріст листків; викликає затримку мітозу. Усі явища знімаються підвищеною концентрацією СО2. Обробка етиленом індукує утворення коренів на стеблі; викликає епінастію (опускання) листків. У багатьох видів прискорює проростання пилку, насіння, бульб, цибулин. Та найефективнішим проявом дії етилену є регуляція процесів дозрівання плодів.
Дозрівання плодів - досить складний процес, який супроводжується посиленням інтенсивності дихання, розпадом багатьох сполук, розм'якшенням тканин, зміною їх забарвлення, запаху тощо. Встановлено, що плоди в період дозрівання синтезують етилен, який і регулює всі ці процеси. Етилен підвищує проникність клітинних мембран, що забезпечує можливість контакту ферментів та субстратів, підвищує інтенсивність синтезу білків, необхідних для процесу дозрівання.
Етилен — летючий фітогормон, тому може впливати на ріст сусідніх рослин. Гормон гальмує полярний транспорт ауксину, і саме з цим пов'язують здатність його посилювати процеси старіння, опадання листків та плодів, усувати апікальне домінування.
Роль його в опаданні листків, плодів пояснюється тим, що етилен спричинює розростання клітин розмежувальної (відокремлювальної) зони в основі черешка чи квітконіжки. Ці клітини набувають здатності синтезувати та виділяти ферменти, які руйнують клітинні стінки, — целюлазу, пектиназу. Ферменти атакують клітинні стінки, руйнують їх, рухаючись по клітинах розмежувальної зони. Зв'язок між клітинами переривається, що призводить на кінцевому етапі до опадання листя.
Різко зростає кількість етилену при стресах і пошкодженнях тканин. Вважають, що етилен може виконувати роль тригера — пускового механізму адаптивних реакцій при дії багатьох стресових факторів, наприклад при дефіциті кисню, інфекційних хворобах рослин тощо.
Є дані, що етилен відіграє важливу роль у детермінації статі у дводольних рослин. Так, у гарбузових він збільшує кількість жіночих квіток. На огірках було виявлено, що жіночі бруньки виділяють значно більше етилену, ніж чоловічі. Таким чином, етилен гальмує ріст, прискорює старіння клітин, дозрівання і опадання плодів, формування розмежувального шару в черешках листків і плодоніжках, є гормоном стресу, бере участь у детермінації статі, прискорює проростання пилку та насіння.
Механізм дії етилену на молекулярному рівні маловідомий. Однак встановлено, що він стимулює синтез раніше зазначених ферментів, а також специфічно може зв'язуватись з білком-рецептором мембран ЕР та апарату Гольджі, активуючи таким чином утворення певних ферментів. Ймовірно, що етилен впливає на стан цитоскелету у клітин.
Один з можливих механізмів дії етилену такий. Рецептор етилену - гістидинкіназа - складається з двох трансмембранних білків: сенсорного (власне гістидинкіназа) і регуляторного. Етилен поєднується з рецептором, який локалізується в плазмалемі, при наявності іонів міді. У разі приєднання гормону відбувається інактивація рецептора. За відсутності етилену рецептор проявляє функції гістидинкінази, яка активує білок, що є головним у передачі сигналу. Наступним елементом сигнального ланцюга є ядерний білок (транскрипційний фактор), який поєднується з промотором гена-мішені та активує його експресію.
У виробничих умовах використовують препарат етефон, який під час обробки ним рослин розкладається з виділенням етилену. Синтетичні препарати використовуються для синхронізації цвітіння (наприклад, ананасів) та прискорення післязбирального дозрівання плодів.
Електрофізіологічна регуляція
Подразливість — одна з найважливіших властивостей живих систем, яка лежить в основі різноманітних рухів та інших проявів життєдіяльності рослин. На відміну від тваринних організмів, у рослин відсутня нервова система, тому функція збудження притаманна для всіх її елементів. Інтегральним показником подразливості у рослин є біоелектричні потенціали. За останні роки встановлено, що всі рослини за певних умов здатні генерувати потенціал дії (ПД). Як виявилося, у координації функціональної активності та в процесах морфогенезу важливе значення мають електрофізіологічні взаємодії клітин, тканин, органів.
Електротонічні поля та струми
Отже, електрофізіологічна регуляція бере участь у керуванні діяльністю та взаємоузгодженні функціональних процесів, що відбуваються в різний час у рослинному організмі. Як це відбувається?
Встановлено, що вегетуюча верхівка вищих чи нижчих рослин відносно більш базальної частини зазвичай заряджена позитивно; центр стебла електропозитивний щодо зовнішньої поверхні; верхівка колеоптиля від'ємна стосовно основи; кінчик кореня позитивний щодо іншої частини кореня; наземна частина рослини звичайно електропозитивна відносно кореня. Між ними виникає струм порядку 0,1-0,4 мкА.
Електропозитивація у рослин у більшості випадків пов'язана з діяльністю Н+- помпи і характерна для ділянок із високою метаболічною активністю.
Як бачимо, між різними частинами рослинного організму існують стаціонарні, повільнозмінні різниці потенціалів, а також потенціали дії. Іншими словами, між ними існують електротонічні поля та струми. Такі види електричної активності й лежать в основі електрофізіологічної регуляції. Зміни в іонних потоках, що виникають за дії різноманітних факторів, спричинюють виникнення нових величин мембранних потенціалів. Існує думка, що під впливом електротонічних полів у клітинних мембранах відбувається латеральне переміщення заряджених ліпопротеїнових комплексів, які виконують різноманітні функції. Отже, в результаті зміни мікроструктури електричних полів у тканинах може бути перерозподіл рухомих білкових компонентів у мембранах, що буде зумовлювати новий фізіологічний стан клітини.
Таким чином, між різними частинами рослинного організму існують відносно стаціонарні (повільно-змінні) різниці потенціалів (РП) - електротонічні поля та струми. Усі зовнішні й внутрішні зміни призводять до зміни величин мембранного потенціалу (МП) клітин або РП між різними частинами рослинного організму. Зміна електротонічних струмів зумовлює в рослин зміни у ростових реакціях, морфогенезі, рухах.
Потенціал дії
На плазмалемі рослинних клітин у результаті діяльності іонних помп виникає МП. Він може змінюватися в залежності від стану зовнішніх та внутрішніх умов. У збудливих клітин місцеве зниження МП до критичного рівня спричинює дальше швидке його падіння (фаза деполяризації), після чого МП повертається до величини, близької до вихідного значення. Так виникає потенціал дії (ПД), або спайк, який має здатність поширюватися.
Остергаут у 30-х роках уперше виявив це явище в харових водоростей, а пізніше його зафіксували також у венериної мухоловки (25 см/с), у мімози (4 см/с). У більшості рослин ПД поширюється зі швидкістю 0,08-0,5 см/с. Він рухається по плазмалемі і плазмодесмам паренхімних клітин флоеми і протоксилеми провідних пучків.
Механізм спайка:
- фаза деполяризації пов'язана з входом у клітини іонів Са2+ і виходом Cl-;
- реполяризація пояснюється виходом К+; Н+- і К+-насос відновлює рівновагу.
Таким чином, у рослин існує швидкий електричний зв'язок, хоча об'єм інформації, яку він передає, незначний.
Сигнальна роль ПД виявляється при проростанні пилку, при різкій зміні умов існування в зоні коренів, пагонів. Прикладом може бути потрапляння пилку на приймочку кукурудзи, в результаті продукується ПД, який надходить до зав'язі та спричинює там відповідні біохімічні перетворення. Зміна хімічного складу ґрунту в зоні кореня продукує одинокий імпульс, який, сягаючи листка, змінює його функції. Подразнення верхівки пагона спричинює ПД, який прискорює поглинання мінеральних іонів. Прилипання комах до листка росянки викликає ПД, який змінює рухову й секреторну діяльність даного органа.