Основи еволюції - Корж О.П. - 2006
Частина III.ШЛЯХИ РОЗВИТКУ ЖИТТЯ
Розділ 18. Розвиток нижчих організмів
18.3.Три ароморфози протерозою
Протерозойську еру можна вважати одним із найважливіших етапів загальної еволюції організмів, оскільки в ній відбуваються три глобальні ароморфні перетворення, які мали суттєвий вплив на подальший розвиток життя на Землі. Сталися ці ароморфози практично одночасно, тому розмова про них піде в одному розділі.
Першою надзвичайно важливою подією можна вважати появу еукаріотичної клітини.
Бактерії - найпростіші дрібні організми, які здатні розмножуватися шляхом простого поділу на два. У природі вони займають всі екологічні ніші, що пояснюється значним різноманіттям їх біохімічного складу. Вважається, що з переходом від відновлювальної атмосфери до окиснювальної (накопичення О2), первинні анаеробні форми перестають відігравати помітну роль у розвитку біосфери (на відміну від аеробних). Вільний кисень, який взаємодіє з більшістю компонентів цитоплазми, був токсичним для анаеробів. Проте завдяки його високій реакційній здатності аероби отримують значно ефективніше джерело хімічної енергії.
Подолання цієї проблеми могло відбутися двома шляхами - перехід до анаеробних умов існування або формування аеробних організмів. Нині процес дихання виступає основним окиснювальним шляхом переважної більшості організмів. Одним із найважливіших виявів пристосування до аеробних умов існування стає формування еукаріотичної клітини, яка набуває більшої стабільності порівняно з прокаріотами.
Існують дві принципово протилежні теорії походження еукаріотичних організмів: симбіогенезу та несимбіотичного зародження — шляхом виокремлення клітинних структур із протоплазми прокаріотичної клітини. Теорія симбіогенезу, яку запропонував К.С. Мережковський у 1905 році, є найбільш логічним поясненням виникнення такого складного утворення, як еукаріотична клітина.
Припускається, що еукаріоти сформувалися внаслідок симбіозу анаеробної прокаріотичної клітини амебоїдного типу та аеробних бактерій (останні дають початок мітохондріям). Без мітохо- ндрій клітини тварин та грибів були б облігатними анаеробами і цілком залежали б від малоефективного процесу гліколізу. З набуттям мітохондрій плазматична мембрана клітини передає їм енергетичну функцію, а сама починає виконувати нові - контроль проходження іонів тощо, чого немає в прокаріотів. Саме поява мітохондрій надає клітинам ефективне джерело енергії.
Можливість симбіотичного походження мітохондрій підтверджується наявністю деяких сучасних анаеробних форм (амеби на зразок Pelomixa palustris), які не мають своїх мітохондрій, а окиснювальний метаболізм здійснюють завдяки симбіозу з аеробними бактеріями. Найважливіший органоїд аеробної клітини - ядро також утворився симбіотичним шляхом (йому дає початок джгутикова прокаріотична клітина).
Наступним ароморфним перетворенням слід вважати появу фотосинтезу та формування рослин як особливої групи організмів.
Хлоропласти здійснюють фотосинтез за тим самим принципом, що й прокаріоти (ціанобактерії), а з деякими їх видами існує морфологічна схожість. Хлоропласти, як і ціанобактерії, розмножуються поділом, а нуклеотидна послідовність ДНК майже повністю гомологічна певним ділянкам їх хромосом. Можливо, що хлоропласти мали спільного предка з ціанобактеріями і утворилися з прокаріот, які були поглинуті еукаріотичними клітинами.
Мітохондрії та хлоропласти мають спільні риси з сучасними аеробними бактеріями та ціанобактеріями, але є між ними й чимало відмінностей. Це пов'язано з тим, що вони пройшли довгий шлях суттєвих еволюційних перетворень і стали досить залежними від клітин-хазяїв.
Згідно з теорією несимбіотичного походження еукаріотичної клітини, утворення органел (ядро, мітохондрії тощо) відбувалося шляхом інвагінації клітинної стінки з подальшим її виокремленням та ускладненням.
Оскільки генетична інформація еукаріотичної клітини зосереджена в ядрі й замкнена його оболонкою, то це унеможливлює прямий поділ самої клітини. Тому виникає необхідність в усуненні оболонки ядра і в чіткому механізмі розподілу генетичної інформації між дочірніми клітинами. Таким механізмом стає мітоз.
Новим етапом еволюції еукаріотичних організмів є поява статевого розмноження, яке пов'язане з виникненням мейозу. Під час його здійснення відбувається перемішування та рекомбінація геномів (комбінативна мінливість), унаслідок чого з'являються особини з новими генотипами. Процеси ці певним чином позначаються й на самому ході подальшої еволюції - відбувається прискорене формування нових ознак.
Вищі рослини (крім мохоподібних) та тварини більшу частину життєвого циклу проводять в диплоїдній фазі, а гаплоїдна фаза в них дуже коротка. Процес еволюції сприяв статевому розмноженню, оскільки випадкова генетична рекомбінація підвищує шанси організмів на виживання в дуже мінливому навколишньому середовищі. Статевий процес також необхідний для підтримання диплоїдності та створення умов швидкого вироблення нових генів у рослин і тварин. Це значно прискорює загальний плин еволюції.
Одноклітинні організми відносно добре пристосовані до навколишнього середовища — деякі з них мають досить складну форму. Проте багатством життєвих форм та структурою утворених ними екосистем вони значно поступаються багатоклітинним організмам.
Поява багатоклітинності (третє ароморфне перетворення протерозою) призвела до значного урізноманітнення організмів на планеті, що підвищило «тиск життя» і, як наслідок, прискорило загальний хід еволюції.
Існує кілька теорій походження багатоклітинних організмів, з яких найбільш відомі дві — теорія гастреї та теорія фагоцители.
Теорію гастреї запропонував німецький учений Е. Геккель у 1872 році (теорія походження багатоклітинних тварин від гіпотетичного первісного організму - гастреї). Згідно з його поглядами, гастрея нагадувала одну зі стадій зародкового розвитку багатоклітинних організмів — гаструлу і мала вигляд видовженого мішкоподібного тіла з двошаровою стінкою та центральною порожниною, яка відкривалася в навколишнє середовище бластопором (первинним ротом). Гастрея виникла з бластеї (одношаровий кулястий організм) шляхом інвагінації. Ця теорія недоказова, оскільки не існує достатніх підстав вважати інвагінацію первинним способом гаструляції. За цією самою теорією виходить, що такі первиннороті тварини, як плоскі та круглі черв'яки, у яких немає целома, втратили його вторинно, але це не відповідає дійсності.
Теорію фагоцители (походження багатоклітинних організмів від джгутикових колоніальних форм) запропонував І.І. Мечников у 1877 році. Згідно з його уявленнями, еволюція предків багатоклітинних, тобто подібних до бластули колоніальних джгутиконосців, відбувалася шляхом диференціації клітин на локомоторні й травні, а також зростання їх взаємозалежності: травні клітини втрачали джгутики, переходячи до амебоїдного стану, занурювалися в порожнину і утворювали внутрішню масу клітин - фагоцитобласт; джгутикові ж клітини зберігали свою локомоторну функцію. Так сформувався спільний двошаровий предок багатоклітинних тварин - фагоцитела (рис. 18.3).
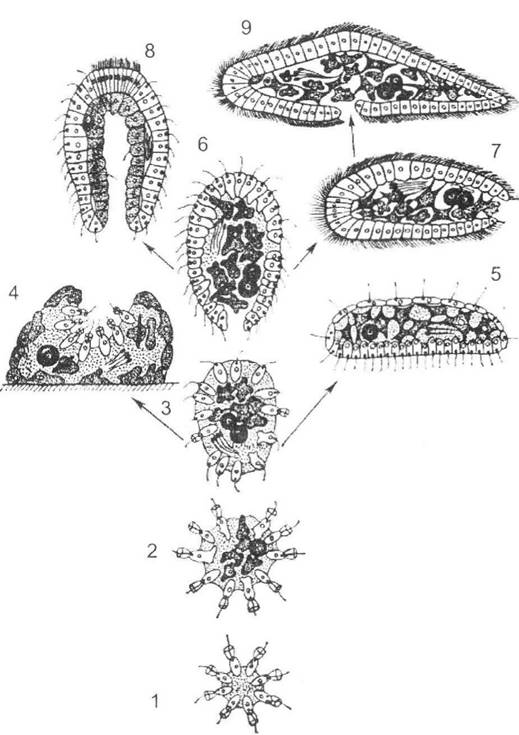
Рис. 18.3. Найважливіші передбачувані стадії раннього філогенезу багатоклітинних організмів (за Ю.І. Полянським, 1987):
1 - колонія джгутиконосців; 2 - міграція частини клітин всередину; 3 - фагоцитела; 4 - губка; 5 - трихоплакс; 6 - пізня фагоцитела з ротовим отвором; 7 - первинна білатеральна тварина; 8 - спільний предок кишковопорожнинних; 9 - первинний плоский черв'як
Згідно з сучасними поглядами, багатоклітинні організми походять з колоніальних форм. Особливо добре ймовірність такого розвитку простежується при вивченні зелених джгутиконосців, серед яких є одноклітинні, колоніальні та багатоклітинні організми. Із колоніальних найбільший інтерес викликають вольвоксові, в яких вже розподіляються функції між клітинами колонії: репродуктивна властива лише незначній кількості клітин, а всі інші втратили свою здатність до розмноження й до самостійного існування. Таким чином, у клітин вольвокса в примітивній формі виявляються дві суттєві риси всіх багатоклітинних організмів:
1) спеціалізація і 2) кооперування. Це призводить до утворення єдиного координованого організму, який має більше можливостей, ніж його будь-яка складова частина.
А.В. Іванов передбачає, що початок багатоклітинним могли дати незабарвлені колоніальні джгутиконосці. Відкриття наприкінці XIX століття трихоплакса дозволило заповнити прогалину між одно- та багатоклітинними організмами. Наближеність цієї тварини за будовою до фагоцители підтверджує правильність поглядів І.І. Мечникова.